|
|
|
Bactéries et virus
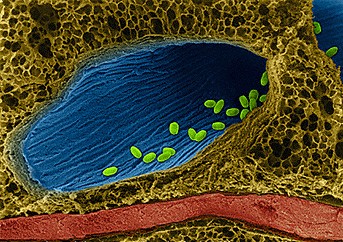
Des bactéries aux deux visages (II) La plupart des bactéries vivent dans l'intestin et en particulier le colon des êtres humains et des animaux, dans la bouche, sur la peau, sur le sol ou dans les aliments et sont inoffensives. Ce sont des agents biohimiques vecteurs de nombreuses transformations (ferments lactiques, etc) mais ils peuvent occasionnellement et par manque d’hygiène devenir infectieux. On peut dire la même chose de certains acariens qui vivent également sur notre peau comme le fameux Demodex folliculorum qui vit notamment dans les pores de notre visage (et oui). Décrivons trois bactéries bien connues parmi les 75000 espèces connues sur les millions existantes. Le fameux pneumocoque (Streptococcus pneumoniae) est une bactérie commune présente chez 5 à 25% de la population dans la bouche, le nez et le pharynx. Tant qu'il reste cantonné à ces endroits la personne peut rester en bonne santé. En revanche, s'il descend vers les voies respiratoires, il peut provoquer une pneumonie. S'il remonte vers les oreilles, il peut provoquer une otite chez l'enfant. S'il infecte les muqueuses nasales, il peut provoquer des sinusites chez l'adulte. Enfin, s'il migre dans la circulation sanguine, il peut provoquer une méningite, une septicémie ou une bactériémie. Toutes variétés confondues, les pneumocoques sont porteurs de 90 types différents d''antigènes. La plupart des vaccins destinés aux adultes sont constitués d'un mélange des 23 antigènes principaux (vaccin 23-valent). Il existe également des vaccins conjugués qui protègent les enfants (entre 2 mois et 2 ans) contre 10 ou 13 souches bactériennes. Le streptocoque est une bactérie arrondie que nous portons tous. On la retrouve dans la transpiration notamment (sous les aisselles, sur les bras, etc). Elle est généralement inoffensive et tous les sportifs en pleine action vivent très bien avec. Toutefois, certaines souches dites à streptocoques A (Streptococcus pyogenes) sont porteuses de germes pathogènes à l'origine d'infections bénignes et non invasives (angine, impétigo, inflammation, etc) ou sévères et invasives (syndrome de choc toxique, bactériémie, infection cutanée nécrosante, méningite, etc). En revanche, les infections à streptocoques B (Streptococcus agalactiae) provoquent des infections graves et parfois mortelles. En de très rares occasions le streptocoque peut subir des mutations et devient un véritable tueur. En effet, s'infiltrant dans le bras ou la jambe par exemple par le biais d'une petite plaie, il s'attaque aux chairs au point de les détruire, les zones infectées progressant jusqu'à 2.5 cm par heure ! Ce streptocoque mutant est capable de tuer un homme de manière fulgurante en l'espace de 24 heures si la zone infectée n'est pas extraite ou le membre amputé. Heureusement on recence très peu de cas de cette gravité.
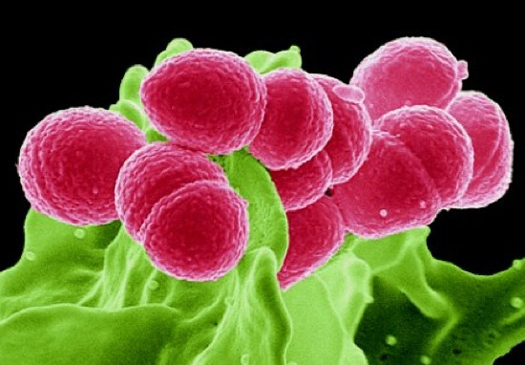
Une autre espèce à deux visages est le staphylocoque doré (S.aueus). C'est une bactérie opportuniste qui devient pathogène au contact des toxines et de facteurs virulents. Elle devient alors très invasive et peut devenir dangereuse. Ce n'est donc pas sans raison qu'elle a mauvaise réputation et qu'elle fait l'objet de beaucoup d'études. S.aureus est présent en permanence dans 20% de la population et jusqu'à 60% de la population de manière temporaire. On peut parfois le trouver dans les cavités antérieures des fosses nasales, dans la gorge et le tube digestif notamment. Tant qu'il y demeure il peut rester inoffensif. Mais si suite à une blessure ou un autre agent pathogène (l'ingestion de nourriture périmée par exemple contenant des entérotoxines produites par S.aureus) il se multiplie et se dissémine et monte vers le cerveau par exemple ou descend vers le tube digestif, les poumons, le périnée ou infecte la peau, il peut devenir toxique et provoquer de graves infections. Raison pour laquelle sa présence dans les hôpitaux est toujours redoutée. Vu ses effets, il est fortement déconseillé de s'arracher les poils du nez par exemple car en cas de blessure ou d'infection cela peut offrir à cette bactérie une voie d'accès directe au sang, aux méninges, aux poumons ou aux os et entraîner de sérieuses infections, voire dans le cas extrême entraîner la mort si l'infection est généralisée (le cas d'une septicémie) et si la personne n'est pas prise en charge rapidement. Les infections qu'il provoque (nosocomiale, panari, septicémie, etc) peuvent normalement se soigner avec des vaccins ou des produits antisceptiques et le traitement ne dure que quelques jours. Si les symptômes persistent ou s'aggravent, il faut prendre des antibiotiques à base de β-lactame telle la Pénicilline ou la Méthicilline (il existe une dizaine de familles), et le traitement peut durer 2 semaines. Toutefois, le staphylocoque doré a développé une résistance à ces antibiotiques (la souche SARM ou MRSA) ce qui augmente la difficulté de le combattre avec les traitements traditionnels, ce qui explique parfois le recours à l'hospitalisation. Mais même dans un hôpital a priori à l'abri des microbes, cette bactérie peut se cacher et infecter des personnes saines d'où l'obligation de respecter à la lettre les consignes d'hygiène. Malgré ce titre aux relents de scénario de science-fiction, la résistance des bactéries est une réalité bien concrète et qui devient alarmante. Ne prenons qu'un exemple extrait de l'actualité. Selon le webzine "Wired", chaque année aux Etats-Unis deux millions de personnes contractent des maladies à cause de bactéries et de champignons qui sont devenus résistants à certains antibiotiques; ces organismes ont subi des mutations et les protéines qu'ils portent ne sont plus reconnues par le système immunitaire ou le vaccin censé lutter contre cette souche microbienne. Parmi ces malades, plus de 13% soit 23000 personnes en meurent. Indirectement, ces soins et ces décès augmentent les dépenses en soins médicaux de 20 milliards de dollars. L'Europe est également confrontée au même problème. L'ECDC (Centre Européen de Contrôle des Maladies) estime que 25000 décès par an résultent de la résistance aux antibiotiques. Depuis quelques années, la résistance des bactéries devient un véritable problème de santé au point que l'OMS a recommandé aux médecins d'éviter de prescrire des antibotiques dans la mesure du possible et de lancer le slogan "les antibiotiques, c'est pas automatique" même si cet avis peut déplaire aux patients concernés. En effet, il existe des centaines de formules chimiques pour créer des vaccins et des antibiotiques. Ils ont un spectre plus ou moins large et peuvent donc tuer différents types de bactéries, sauf si elles deviennent résistantes ou multirésistantes. Selon les projections des spécialistes, si on continue à prescrire des antibiotiques pour des infections bénignes, on estime qu'en 2050 la mortalité liée aux microbes pathogènes multirésistants sera supérieure à tout autre risque de mortalité. Bref, on reviendra à la situation sanitaire d'avant l'invention de la Pénicilline où les infections et la gangrène étaient les principales causes de mortalité ! On reviendra sur le sujet à propos de l'élimination des bactéries et des virus. L'une de ces bactéries résistantes est la fameuse Listeria. La Listeria Chaque année en Belgique, l'Institut de Santé Sciensano relève en moyenne 70 à 75 infections à la Listeria ou listérioses. Cette bactérie, Listeria monocytogenes, est la seule espèce de Listeria pathogène pour l'être humain (cf. Radoshevich & Cossart, 2018).
La Listeria est commune dans l'environnement et présente la particularité d'être très robuste. Selon Pascale Cossart de l'Institut Pasteur, "elle résiste aux outils qu'on utilise normalement dans la lutte contre les bactéries. Par exemple, elle pousse à basse température : elle se multiplie à 4 degrés. Elle résiste aussi à de très fortes concentrations salines." Même placée au frigo, la Listeria continue à se développer contrairement à la plupart des autres bactéries et champignons que le froid met en léthargie ou dégrade. La Listeria est donc l'une des rares bactéries qu'on retrouve dans la chaîne du froid où elle continue à se multiplier. D'où l'importance de vérifier les dates de péremption des aliments et en particulier de la charcuterie, du poisson (y compris du saumon en tranches et du Surimi) et des fruits de mer placés au frigo (à domicile ou dans les supermarchés). Heureusement la Listeria succombe à 60°C, raison pour laquelle il n'y a pas une seule bactérie dans le fromage à base de lait pasteurisé ou à pâte pressée cuite (Abondance, Beaufort, Compté, Gruyère, Emmental, etc). Mais on en trouve dans le fromage au lait cru (Brie, Reblochon, Roquefort, Salers, etc) aux côtés de E.coli, des Salmonelles, ... La Listeria est une bactérie dite résistante car elle peut se réfugier à l'intérieur des cellules où le système immunitaire a beaucoup plus difficile de la débusquer et la tuer. Même les macrophages censés phagocyter - absorber et détruire - les corps indésirables, ne la détruisent pas. Pire, la Listeria est capable de franchir les différentes "barrières" de protection sanitaire du corps humain : la barrière intestinale (où elle peut se retrouver dans le sang), la barrière hémato-encéphalique (qui conduit au cerveau) et la barrière féto-placentaire (vers le fœtus chez les femmes enceintes). Quand la Listeria réussit à traverser ces barrières, elle peut donc devenir pathogène mais seulement si le terrain est propice. La Listeria est mortelle dans 20 à 30% des cas. Selon le microbiologiste Jean-François Collet de l'Institut de Duve de l'UCL, une personne en bonne santé ne risque pas grand chose. En fait, il faut qu'une personne saine ingère de la nourriture contaminée contenant au moins un milliard de bactéries pour ressentir des maux de ventre mais sans autre effet. En revanche, une personne fragilisée contaminée par 100 à 10000 bactéries peut développer une maladie potentiellement mortelle car les Listeriae s'engouffreront dans les brèches du système immunitaire, même aux endroits où normalement il n'y a pas de bactéries, comme le cerveau.
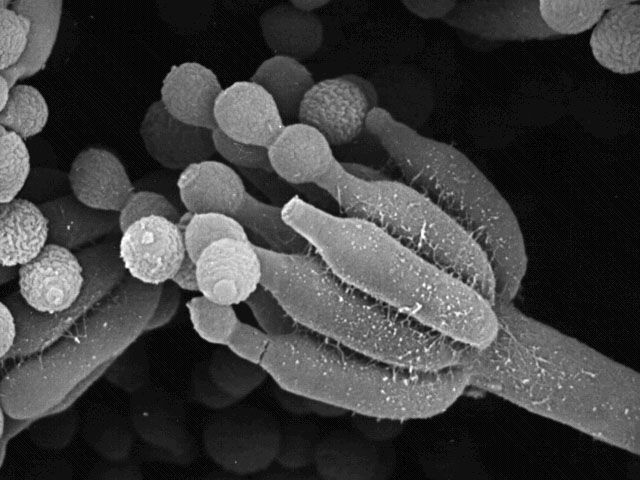
Mais qu'on se rassure. Les maladies liées à la Listeria sont peu fréquentes et il existe des traitements efficaces dont les antibiotiques prescrits à fortes doses et des traitements assez longs qui viennent à bout de la listériose. Il existe également des anticorps spécifiques comme le clone LZH1. Ceci dit, le meilleur remède est l'hygiène : se laver régulièrement les mains, laver son frigo périodiquement, respecter les dates de péremption, bien cuire la nourriture, préférer les aliments non transformés, etc. Comment les bactéries deviennent-elles multirésistantes ? Le transfert horizontal de gènes et la conjugaison bactérienne L'une des facultés de certaines bactéries comme E.coli est leur capacité à compléter leur ADN. Il existe plusieurs mécanismes. Comme on le voit ci-dessous à gauche, E.coli dispose d'un appendice spécialisé appelé "pilus" qui n'est pas un flagelle (utilisé pour les déplacements) mais une sorte de tube flexible qui lui permet de transférer des informations génétiques en dehors du processus de reproduction habituel par mitose (division par scissiparité). Il existe trois types de pilus : le pilus commun qui assure l'adhésion cellulaire et participe à l'activité pathogène de la bactérie, le pilus sexuel qui permet de transférer des informations génétiques et le pilus électroconducteur qui permet de capturer des électrons libres. Celui qui nous intéresse est le pilus sexuel car il permet aux bactéries de recevoir de l'ADN d'un donneur ou de capturer des fragments d'ADN disséminés dans le milieu (par exemple sur un cadavre de bactérie dont le cytoplasme et l'ADN se sont répandus alentour suite à la lyse - destruction - de sa paroi cellulaire). Comme un bras articulé versatile ou une pompe suceuse, la bactérie projète son pilus sexuel exactement là où se trouve un brin d'ADN qu'elle intègre ensuite dans son propre ADN. Ce mécanisme est appelé le transfert horizontal de gènes (par opposition au transfert vertical où le matériel génétique est hérité de l'ancêtre). Quand il s'établit entre deux organismes on parle de conjugaison bactérienne.
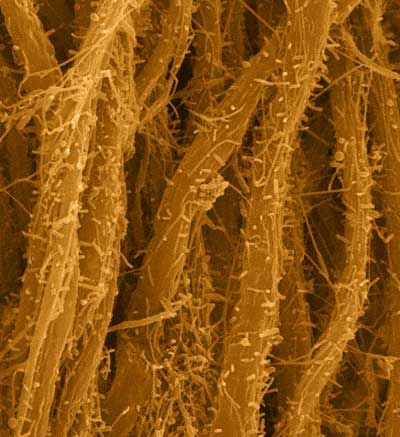
Dans un article publié dans la revue "Nature Microbiology" en 2018, le biologiste Ankur B. Dalia de l'Université d'Indiana et ses collègues ont expliqué ce mécanisme sur base de l'observation in vivo de la bactérie Vibrio cholerae porteuse du choléra en train de capturer de l'ADN comme on le voit ci-dessus à droite. Du fait que les structures bactériennes impliquées sont extrêmement petites (< 0.1 micron), c'est la première fois qu'on observa ce mécanisme pourtant connu depuis des dizaines d'années. Comme l'explique la biologiste Courtney K. Ellison coauteure de l'article, la bactérie projète son pilus sexuel de manière tout à fait ciblée : "C’est comme enfiler une aiguille. La taille du pore de la membrane extérieure [de la bactérie réceptrice] correspond presque à celle d’un brin d'ADN plié en deux, ce que semble avoir attrapé la bactérie. Si le pilus n’était pas là pour la guider, la probabilité que l’ADN atteigne le pore au bon moment et avec le bon angle pour traverser la paroi, est proche de zéro." Le transfert horizontal de gènes représente un moyen important pour les bactéries d'améliorer leur ADN et d'évoluer. Car à force de partager leur ADN ou suite à des mutations (des erreurs de transcriptions lors de leur division), les bactéries peuvent devenir résistantes et même multirésistantes, c'est-à-dire capables de résister à plusieurs antibiotiques. Il est donc important de comprendre ce mécanisme, car plus nous en savons sur la manière dont les bactéries partagent leur ADN, plus les biologistes pourront aisément le contrer. A consulter : EnsemblBacteria Plus de 20000 génomes bactériens en ligne
Les bactéries multirésistantes ou BMR sont 40 fois plus nombreuses qu'à la fin du XXe siècle et le nombre d'épisodes (de personnes infectées) augmente chaque année. Cela signifie qu'en cas d'infection, les bactéries résistantes se multiplient au détriment des souches normales et peuvent, sous certaines conditions, créer une véritable pandémie. Certaines bactéries deviennent même hautement résistantes, pour lesquelles il n'existe aucun remède. C'est le cas de la spasmonelle kentuki, qui sévit surtout en Asie du Sud-Est. Si elle se dissémine et débarque sur un autre continent, les scientifiques n'ont aucun moyen de l'arrêter, preuve que le combat contre les microbes pathogènes est une lutte au quotidien. Ne pas confondre cette forme de résistance avec la résistance naturelle de certaines bactéries aux antibiotiques comme les sp. Borrelia que transportent les tiques infectées vecteurs de la maladie de Lyme ou borréliose. Seuls deux antibiotiques spécifiques peuvent traiter cette maladie bénigne, traitement qui exige donc une analyse de sang préalable. Ceci dit, il existe des thérapies alternatives. On y reviendra à propos des maladies de la peau. La cilagicine, un nouvel antibiotique prometteur Des chercheurs de l'Université Rockefeller de New York ont annoncé en 2022 avoir synthétisé un nouvel antibiotique à l'aide de modèles informatiques de produits génétiques bactériens. Cette substance tue même les bactéries résistantes aux autres antibiotiques. La molécule appelée "cilagicine" a été testée sur des souris et utilise un nouveau mécanisme pour attaquer des bactéries à l'origine d'infections nosocomiales comme le Staphylocoque doré (Staphylococcus aureus Résistant à la Méthicilline ou SARM), le Clostridium difficile et plusieurs autres agents pathogènes mortels (cf. S.F. Brady et al., 2022).
Selon Sean F. Brady, professeur Evnin et auteur correspondant de cette étude, "Ce n'est pas seulement une nouvelle molécule cool, c'est une validation d'une nouvelle approche de la découverte de médicaments. Cette étude est un exemple de biologie computationnelle, de séquençage génétique et de chimie de synthèse réunis pour percer les secrets de l'évolution bactérienne." Reste à présent à réaliser les essais cliniques sur les humains. Si les tests sont concluants, cet antibiotique pourrait donc être disponible d'ici un an ou deux. Les inconvénients de la promiscuité Dans notre vie quotidienne, le problème de la résistance des bactéries apparaît surtout dans l'élevage en batterie, où la densité des animaux est anormalement élevée comparée au milieu naturel (par exemple 1 poule sur la surface d'une feuille A4 !). Le risque d'infection y est élevé d'où l'emploi régulier d'antibiotiques. Mais revers de cette stratégie, elle donne naissance indirectement à des souches multirésistantes. Aussi depuis quelques années, on ne peut plus donner n'importe quels antibiotiques dans les élevages pour justement essayer d'enrayer la prolifération des bactéries résistantes. Ce type d'élevage n'est donc pas une solution très saine et les médias s'en font régulièrement l'écho. C'est ici que la viande Bio et ses dérivés (les oeufs Bio, etc) prennent tout leur intérêt. Nous pouvons citer un autre exemple où la promiscuité facilite le transport des agents infectieux. Tous les parents savent qu'en plaçant leur bébé dans une crèche, les bébés plus fragiles que les autres peuvent contracter des infections. Les soigner en les bombardant d'antibiotiques à titre préventif ou curatif est un réflexe qui peut protéger l'enfant contre des maladies bactériennes comme la scarlatine ou la méningite. Mais mieux vaut d'abord voir comment le système immunitaire de l'enfant peut lutter seul contre cette bactérie avant de lui prescrire des antibiotiques adaptés à son âge.
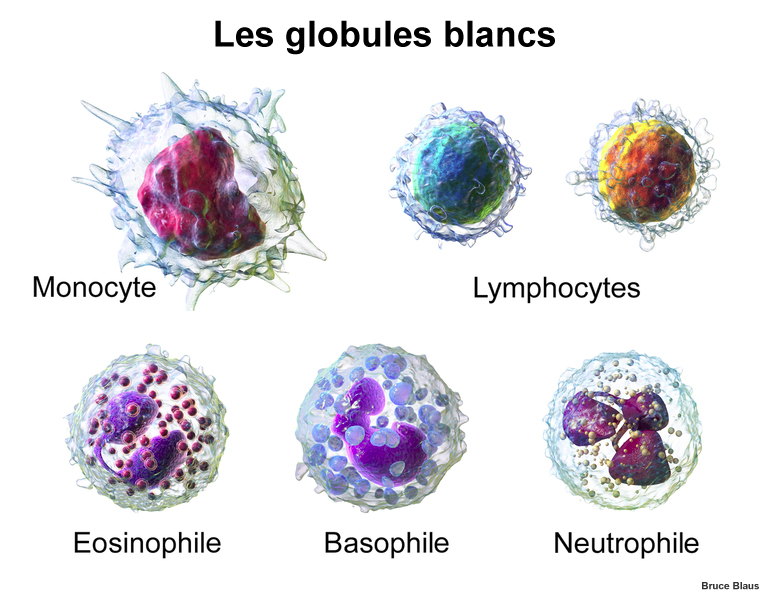
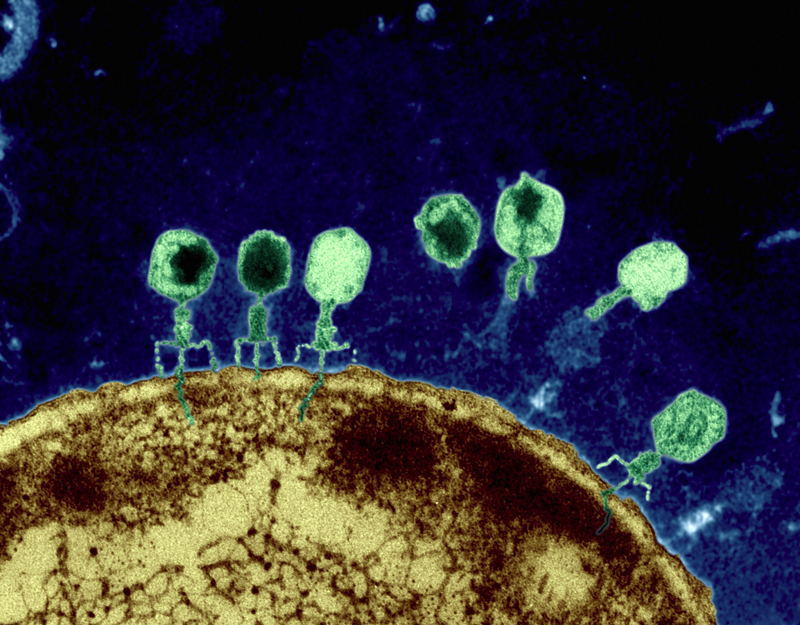
Ici encore, le meilleur traitement est la prévention : isoler le malade, retirer l'enfant sein temporairement de l'établissement et si ce n'est pas possible, de bien se laver tout le corps et en particulier les mains, premiers vecteurs de parasites. Ceci dit, les parents ayant de petits enfants entendront parfois la responsable de leur crèche leur conseiller de laisser leur enfant malade dans l'établissement afin que leurs camarades acquièrent les anticorps pour lutter contre cet agent pathogène. Si effectivement la plupart des autres enfants se porteront bien, certains d'entre eux tomberont malade au grand dam de leurs parents. Ce n'est donc pas un conseil très sage que de laisser un enfant malade parmi des enfants sains. Comment éliminer les bactéries ? L'organisme dispose de deux méthodes pour éliminer les bactéries : les défenses du système immunitaire, c'est-à-dire les différents types de globules blancs notamment que l'organisme peut produire dont les macrophages, des monocytes qui ont la capacité de phagocyter (absorber et détruire) tout corps étranger, et les virus bactériophages. Il s'agit de virus bactériens qui infectent les bactéries en insérant leur matériel génétique dans l’ADN de leur hôte afin de détruire la cellule au cours de la réplication. Ces bactériophages peuvent même lutter contre des bactéries ultra résistantes comme les pseudomonas et les staphylocoques dorés.
Concernant les médicaments, rien ne sert d'utiliser des antiobiotiques à moins que leur spectre d'hôte soit restreint et ciblé sur une bactérie particulière. Si l'infection bactérienne est superficielle, un traitement topique (localisé) suffira à l'éliminer en quelques semaines. Il existe de nombreux remèdes naturels parmi lesquels les lotions antiseptiques à base d’hamamélis qui contient un alcool antibactérien, les produits antifongiques liquides (par ex. l'huile de lavande) ou les crèmes (par ex. Fucicort), le gel désinfectant contenant au moins 60% d'alcool couramment utilisé pour désinfecter les mains. Même le savon est souvent efficace car il permet de dissoudre la membrane lipidique des bactéries (ce qui explique pourquoi les savons ne sont jamais contaminés par des bactéries) et de la plupart des virus enveloppés, d'où l'intérêt de se laver quotidiennement avec du savon. Ce n'est donc pas sans raison que les pandémies sont des situations d'urgence, parfois catastrophiques quand elles entraînent la mort, et qu'on demande à chacun de se laver tous les jours et de se laver les mains plusieurs fois par jour. Nous verrons plus loin que la situation est encore plus dangereuse avec les virus. Pour désinfecter du mobilier et des ustencils, sachant que tous les microbes sont sensibles à l'oxydation, on utilise l'eau de Javel ou l'eau ozonée pour les éradiquer ainsi que les rayonnements ionisants, raison pour laquelle on utilise des stérilisateurs UVC (254 nm ou 222-207 nm) dans tous les lieux médicalisés. L'effet limité de la stérilisation par micro-onde Certaines personnes mettent leur éponge de cuisine usagée au micro-onde en espérant tuer les bactéries par la chaleur. Ce n'est pas une bonne idée. Dans une étude publiée dans la revue "Nature" en 2017, le microbiologiste allemand Markus Egert de l'Université de Furtwangen à Villingen-Schwenningen et ses collègues ont réalisé une expérience qui démontre que cette pratique risque d'être pire que de ne pas nettoyer l'éponge.
Les chercheurs ont testé 14 types différents d'éponges de cuisine collectées auprès de ménages dans et autour de la ville de Villingen-Schwenningen, en Allemagne. Les chercheurs ont coupé ces éponges et prélevé 28 échantillons des faces vertes et jaunes de chacune d'elles. Ils ont ensuite utilisé la technique du séquençage de l'ADN pour déterminer quels types de microbes vivaient sur les éponges et un microscope confocale à balayage laser (FISH-CLSM) pour identifier les endroits où ils se reproduisaient. Le séquençage récolta plus de 220000 séquences d'ADN brut, qui représentaient 9 embranchements, 17 classes, 35 ordres, 73 familles et 118 genres de microbes ! Comme d'autres études l'ont rapporté, les chercheurs ont découvert que les bactéries de la famille des Moraxellaceae dominaient dans les éponges, représentant environ 36% des microbes dans les échantillons. Selon les auteurs, " L'analyse des images montra des densités locales allant jusqu'à 5.4 x 1010 cellules/cm3 et confirma la dominance des gammaprotéobactéries." Comme expliqué précédemment, ces microbes commensaux font partie du microbiome de l'environnement bâti (BE). Ces microbes sont donc très communs et typiques de la peau humaine. Les chercheurs en ont trouvé sur toutes les surfaces de cuisine qui ont tendance à être nettoyées avec des éponges, y compris les surfaces de travail, les étagères des réfrigérateurs, les robinets et les cuisinières. On les trouve également sur le linge sale où ils donnent aux vêtements leur mauvaise odeur. Les chercheurs ont également découvert que les protéobactéries, les bactéroïdes et les actinobactéries étaient des embranchements primaires. Cinq des 10 unités taxonomiques opérationnelles les plus courantes - essentiellement comme des espèces - étaient étroitement liées à des bactéries associées à des maladies modérées. La plupart des donneurs ont admis qu'ils n'avaient pas essayé de stériliser leurs éponges au four à micro-onde. Mais parmi ceux qui l'ont fait, cette méthode n'élimina au maximum que 60% des microbes. Mais les chercheurs ont surtout constaté qu'en essayant de nettoyer une éponge de cuisine usagée, on ne fait qu'offrir plus de place aux microbes pathogènes. En effet, selon les chercheurs, "les bactéries résistantes survivent au processus d'assainissement et recolonisent rapidement les niches libérées jusqu'à atteindre une abondance similaire à celle d'avant le traitement." Dans leurs conclusions, les auteurs soulignent "le rôle des éponges de cuisine en tant que points chauds microbiologiques des BE, avec la capacité de collecter et de propager des bactéries ayant probablement un potentiel pathogène." Conclusion, la meilleure solution, disaient-ils, est de se procurer une nouvelle éponge, de préférence chaque semaine. Ajoutons qu'il est sanitairement plus propre et plus efficace de nettoyer les surfaces de la cuisine y compris les étagères du frigo avec des éponges ou des lingettes à usage unique imbibées d'un produit dégraissant et antibactérien, ce dernier éliminant jusqu'à 99.9% des bactéries. On reviendra en détails sur les moyens pour éliminer les bactéries et virus ainsi que sur le rôle des vaccins. Si le corps humain est parfaitement adapté pour lutter contre la plupart des bactéries, on ne peut pas en dire autant des virus face auxquels le corps humain est dépourvu de défenses. Même si nous pouvons les combattre avec des méthodes de bon sens (isolement, meilleure hygiène, incinération des victimes, etc.), les virus restent les maîtres du monde. C'est l'objet du prochain chapitre. Prochain chapitre |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||