|
|
|
Des bactéries biolumineuses au service de l'homme
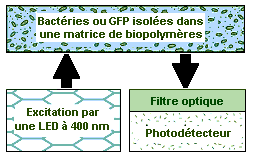
Illustrations du Pr. Gary S. Sayler, CEB Les bioreporters (I) Tout comme les animaux, les microbes sentent les dangers environnementaux avant les êtres humains. Si les premiers utilisent souvent leur "sixième sens" personne jusqu'ici ne peut imaginer le monde sensoriel d'un microbe. Dans ce contexte, comment faire en sorte qu'un microbe communique avec l'homme pour l'avertir par exemple de l'imminence d'un danger ? Le microbiologiste Gary S. Sayler et ses collègues de l'Université de Tennessee ont développé un dispositif utilisant des microprocesseurs capables d'intercepter les signaux émis par des bactéries biolumineuses spécialement modifiées. Ces circuits appelés BBIC ou Circuits Intégrés de Bioreporters Biolumineux sont conçus pour dépister la pollution de l'air. Avec l'appui financier de la NASA, nos chercheurs ont à présent conçu une version pour les vaisseaux spatiaux qui viendrait épauler l'assistant robot autonome, celui qu'on a surnommé le droide S2D2 construit par le centre Ames de la NASA. Gary Sayler qui s'est adjoint les compétences des chercheurs du JPL et du centre spatial Kennedy (NASA-KSC) est parvenu a créer des microbes, qu'il appelle des "bioreporters" émettant une lumière bleue-verte en présence de contaminants comme sur l'image présentée ci-dessus. Couplés à des microprocesseurs sensibles à la lumière, des microluminomètres, ces détecteurs fonctionnent comme n'importe quelle sonde optoélectronique, avec cette particularité unique d'être vivants.
Ces bioreporters sont des cellules microbiennes génétiquement modifiées pour produire un signal mesurable en réponse à un stimulus qui peut être un produit chimique spécifique ou un agent physique présent dans leur environnement. Ils contiennent deux éléments génétiques essentiels, un gène "instigateur" ou promoteur et un gène "reporter" qui rapporte l'information à un système de détection externe. Comme tous les gènes, ils fabriquent des protéines ou participent à la fabrication de l'ADN. Comment fonctionne ce gène bioreporter ? Le gène instigateur s'active (transcrit ou copie son message) quand l'agent auquel il est sensible est présent dans l'environnement de la cellule. Ce gène est normalement lié à d'autres gènes qui sont tous traduits en même temps en protéines pour aider la cellule à combattre par exemple ou s'adapter à l'agent auquel elle a été exposée. Dans le cas du bioreporter, certains de ses gènes (acides aminés) ont été retirés et remplacés avec un gène reporter. En s'activant, le gène instigateur produit une réaction dans le gène reporter. Son activation conduit à la production de protéines reporter qui produisent en fin de compte un certain type de signal, en l'occurrence un signal biolumineux. De cette manière, en présence d'un signal lumineux, le microprocesseur indique que le bioreporter a détecté un agent suspect dans son environnement. Les BBIC, des reporters géniques
Les BBIC, explique Sayler, représentent une manière peu coûteuse et exigeant peu d'énergie pour détecter des polluants. Ils sont minuscules. Comme l'indique la photographie présentée à droite, chaque BBIC mesure environ 2 mm de côté et le dispositif entier, y compris sa source d'énergie, sera probablement de la taille d'une boîte d'allumettes et il pourra surveiller son environnement sans interruption. La NASA s'intéresse à la détection des contaminants car les vaisseaux spatiaux habités sont des lieux de vie scellés où toute pollution peut être mortelle. Les gaz invisibles et inodores libérés par les expériences scientifiques ou les toxines produites par des souches mutantes et autres biofilms peuvent s'accumuler dans les cabines pressurisées et poser un risque pour la santé des astronautes. Les BBIC peuvent être utilisés pour détecter presque n'importe quoi : l'ammoniaque, le cadmium, le chrome, le cobalt, le cuivre, le mercure, le zinc, les protéines, les PCB, les ultrasons, le rayonnement ultraviolet... la liste est longue. Plusieurs types de gènes reporters sont déjà disponibles pour élaborer ces types de détecteurs et les signaux qu'ils produisent peuvent habituellement être classés en différentes catégories : colorimétriques, fluorescents, lumineux, chimiolumineux ou électrochimiques. Bien que chacun fonctionne différemment, leur produit final reste toujours le même : c'est un signal mesurable et proportionnel à la concentration de l'agent polluant. Quelquefois le signal biolumineux émis par la protéine se produit uniquement lorsqu'un substrat additionnel est ajouté à la réaction biologique (par ex. luxAB, luc et Aequorin). Pour d'autres bioreporters, le signal doit être activé par une source lumineuse extérieure (protéines GFP et UMT, voir plus bas). Pour d'autres, moins nombreux, le signal individuel est complètement induit, sans substrat exogène ni activation externe (luxCDABE, voir plus bas). Les gènes biolumineux Voyons en bref certains types de gènes reporters biolumineux et comment peut-on les utiliser. Luciferase bactérien (lux) : La luciferase est un nom générique donné à une enzyme qui catalyse une réaction lumineuse. On peut la trouver dans les bactéries, les algues, les mycètes, les méduses, les insectes, la crevette et le calmar. La lumière qu'elle produit dans ces organismes est bien connue, c'est le phénomène de bioluminescence qu'affectionne beaucoup de créatures abyssales Dans les bactéries, les gènes responsables de la réaction lumineuse (les gènes lux) ont été isolés et employés intensivement dans la fabrication des bioreporters qui émettent une lumière bleu-verte avec une intensité maximum à 490 nm (cyan). Trois variantes de lux existent, l'une fonctionnant en dessous de 30ºC, l'autre à moins de 37ºC et la troisième en dessous de 45ºC. Le système génétique de lux se compose de cinq gènes, luxA, luxB, luxC, luxD et luxE. Selon la combinaison de ces gènes, plusieurs types différents de bioreporters biolumineux peuvent être élaborés.
Bioreporters luxAB : les bioreporters luxAB contiennent uniquement les gènes luxA et luxB qui, ensemble, produisent un léger signal biolumineux. Toutefois, pour accomplir entièrement la réaction lumineuse, un substrat doit être fourni à la cellule. Pratiquement, on le réalise en ajoutant un produit chimique à une certaine phase de la réaction biologique. De nombreux bioreporters luxAB ont été élaborés et incorporés dans le matériel génétique des bactéries, des levures, des insectes, des nématodes, des plantes ainsi que dans des cellules de mammifères. La liste présentée ci-dessous reprend les produits chimiques et les agents biologiques capables d'être détectés par les bioreporters disposant de gènes luxAB.
Bioreporters luxCDABE : au lieu de contenir uniquement les gènes luxA et luxB, chaque bioreporter peut contenir cinq gènes lux, le système résultant n'exigeant aucun apport de substrat ni d'excitation par une source lumineuse extérieure. Ainsi, dans cet essai biologique, le bioreporter est simplement exposé à un agent cible et on observe une augmentation quantitative du signal biolumineux dans un délai souvent inférieur à une heure. En raison de la rapidité de leur réaction et de la facilité d'utilisation, les bioreporters luxCDABE sont extrêmement attrayants. Ils sont donc incorporés à plusieurs méthodes de détection qui s'étendent de la détection des contaminants environnementaux à la surveillance en temps réel des infections par des microbes pathogènes chez les souris vivantes. La table ci-contre illustre les applications les plus répandues des bioreporters luxCDABE ainsi que leur temps de réaction et leur concentration. Bioreporters lux non spécifiques : des bioreporters lux non spécifiques sont employés pour détecter des toxines chimiques. Ils sont habituellement conçus de manière à produire un signal biolumineux de manière continue. Lors de l'exposition à une toxine chimique, soit la cellule meurt soit son activité métabolique est retardée, conduisant à une diminution de l'intensité biolumineuse. Leur application la plus familière est l'analyse Microtox dans laquelle, suivant une exposition courte à plusieurs concentrations de l'échantillon, le signal biolumineux affaibli peut être corrélé avec les niveaux relatifs de toxicité. Luciferase de luciole (Luc) : la luciferase de luciole catalyse une réaction qui produit la lumière visible dans la gamme de 550-575 nm. D'autres variétés de luciferase de coléoptère produisent une lumière proche de 595 nm. Ces deux luciferases exigent l'addition d'un substrat exogène (luciferine) afin que la réaction lumineuse se produire. De nombreux biorécepteurs Luc ont été élaborés pour détecter des composés inorganiques et organiques environnementaux. Toutefois, leurs applications les plus prometteuses se trouvent probablement dans le domaine du diagnostic médical. L'insertion des gènes de luc dans une lignée cellulaire épithéliale (carcinome) permet par exemple de diagnostiquer des cellules tumorales chez une souris vivante en balayant simplement la zone infectée avec une caméra CCD. Ce système permet un traitement chimiothérapeutique rapide et surveillé en temps réel. Dans un autre exemple, les gènes de luc ont été insérés dans des lignées de cellules cancéreuses du sein afin de développer un test biologique pouvant détecter et mesurer les substances ayant une activité estrogénique (liée à la grossesse) et anti-estrogénique potentielle. Aequorine : l'aequorine est une photoprotéine d'isolement que l'on trouve dans les méduses biolumineuses Aequorea victoria. Lors de l'apport d'ions de calcium (Ca++) et de coélentérazine, une réaction se produit dont le résultat est la génération d'une lumière bleue entre 460-470 nm. L'aequorine a été incorporée aux lignées humaines de cellules pour détecter des bactéries et des virus pathogènes dans ce qu'on appelle l'analyse "jaune canari" (des risques d'antigènes). intracellulaires qui libèrent des ions de calcium à l'intérieur de la cellule.
Les cellules sont génétiquement modifiées pour produire l'aequorine. Lors de l'exposition aux antigènes de différents microbes pathogènes, les cellules de recombinaison émettent de la lumière en raison de l'activation d'une cascade de signaux Protéine fluorescente verte (GFP) : La protéine fluorescente verte (GFP) est également une photoprotéine d'isolement extraite des méduses Aequorea Victoria. Des variantes ont été également isolées dans des Renilla reniformis ou pensée de mer (anthozoaire). La GFP, comme l'aequorine, produit un signal fluorescent bleu, mais sans apport de substrat exogène. Tout ce qui est exigé est une source de la lumière UV pour activer les propriétés fluorescentes de la photoprotéine. Ces capacités d'autofluorescence rendent la GFP très attrayante dans les analyses de biosensing puisqu'elle peut être utilisée dans des lignées de cellules et en temps réel pour surveiller les cellules intactes et vivantes. De plus, il est possible de modifier la GFP pour qu'elle produise des émissions biolumineuses différentes du bleu (cyan, rouge ou jaune) permettant de l'utiliser comme détecteur polyvalent sur plusieurs cibles différentes. Comme les gènes luxAB, la GFP a été intensivement utilisée pour élaborer des bioreporters dans quantités d'organismes, des bactéries aux cellules des mammifères. Le tableau présenté ci-dessous énumère quelques exemples des applications de la GFP dans des systèmes cellulaires de mammifères où son utilisation a révolutionné ce que nous savons sur la dynamique des protéines cytoplasmiques, cytosquelettiques et organellaires ainsi que leurs interactions intracellulaires.
Uroporphyrinogène (Urogène) III Méthyltransférase (UMT) : La protéine UMT catalyse une réaction qui rapporte deux produits fluorescents produisant une lumière rouge-orange dans la gamme 590-770 nm lorsqu'elle est illuminée sous les UV. Comme la GFP, cette protéine ne demande aucun apport de substrat exogène. L'UMT a été employée comme bioreporter dans la sélection des plasmides de recombinaison, comme marqueur pour la transcription de gènes chez les bactéries, les levures et les cellules de mammifères ainsi que pour la détection des sels toxiques tels que l'arsénite et l'antimonite. Deuxième partie Détection de la signature optique
|





{kind=link}