|
|
|
Le rôle des virus dans l'évolution
Des traces de virus dans notre ADN (I) Beaucoup de biologistes et de virologues parmi d'autres chercheurs constatent que la théorie de Darwin est incapable d'expliquer l'évolution des espèces autrement qu'en faisant appel au principe philosophique de la "sélection naturelle". On parle d'adaptation, de lutte pour la survie, mais personne ne peut prouver le phénomène en mettant le doigt sur le mécanisme biologique ou chimique ou sur l'entité ayant déclenché ce phénomène (mutation ou autre), encore moins prédire cette évolution si ce n'est à travers l'expression plus ou moins prolongée des gènes. Même les modèles néo-Darwiniens de l'évolution insistant sur les mutations ponctuelles et les recombinaisons sexuelles des gènes de l'organisme hôte (celui abritant le parasite) ne peuvent expliquer l'évolution des espèces. Cherchant plutôt une explication rationnelle à l'évolution des espèces, plusieurs équipes de virologues ont recherché les traces originelles de ce phénomène dans le patrimoine génétique des organismes et notamment chez les bactéries[1] et virus, sachant qu'il s'agit des organismes s'adaptant le plus facilement.
Pourquoi étudier les gènes ? Car depuis les travaux d'Oswald Avery (1944) et de sa démonstration par Al Hershey (1946), nous savons que la molécule d'ADN est porteuse d'informations héréditaires. Toutefois, ce n'est que récemment que le génome humain fut séquencé et cartographié, en particulier grâce aux projets HUGO (1990-2003), 1000 Genomes Project concentré sur les variations génomiques (2008-2015) et Consortium Telomere to Telomere (T2T) qui compléta les 8% de séquences manquantes (2013-2022). Grâce à ce séquençage intégral du génome humain, nous avons découvert que notre ADN contient principalement des séquences ne jouant a priori aucun rôle dans l'expression des gènes, le fameux ADN "de rebut". Plus étonnant encore, des milliers de séquences contiennent des traces de virus inactifs. On estime qu'entre 40 et 80% du génome humain seraient liés à d'anciennes contaminations virales mais dont les vecteurs ont pour ainsi dire eu l'intelligence ou en tout cas l'opportunité de collaborer avec les cellules et les bactéries pour le plus grand bien de l'organisme. Que font-elles là et quel fut leur rôle le cas échéant ? Pour l'essentiel, c'est encore la grande inconnue. On estime que sur Terre il existerait 1031 virus, principalement des phages (des virus infectant les bactéries) qui évoluent dans les océans auxquels il faut ajouter les 800 millions de virus qui tombent chaque jour sur chaque mètre carré de la planète ! On y reviendra. dans l'article Des milliards de microbes nous tombent du ciel. Cela représente un nombre plus élevé que le nombre estimé d'étoiles dans l'univers visible (1024) ! On peut légitimement se demander quel est le rôle de ces virus qui ont colonisé tous les milieux, des roches et des océans les plus profonds au sommet de l'atmosphère et tous les organismes vivants. Logiquement, il est impossible qu'une telle population ne joue pas un rôle significatif dans l'évolution des organismes, soit dans leur développement soit par les interaction qu'elles créent (parasitisme ou symbiose). Et de fait, les observations prouvent que les virus ont joué et jouent encore un rôle actif dans l'évolution des organismes. C'est en étudiant les séquences de l'ADN de rebut du génome humain et en les comparant à l'ADN viral ainsi qu'à celui des bactéries que des virologues ont émis l'hypothèse que les virus seraient en fait les véritables moteurs de l'évolution. En effet, les virus pourraient avoir légué à la postérité des mutations ponctuelles et seraient à l'origine de divergences biologiques majeures et de nouveaux ordres, tels que le développement du noyau eucaryote, le mécanisme de réplication eucaryote et la tolérance immunitaire[2] adaptative. Mais ce postulat a de lourdes conséquences : il signifie d'une part que la théorie de l'évolution de Darwin, même dans sa version moderne, serait fausse - ce que la plupart des observations refutent - et d'autre part que les virus ne seraient pas tous dangereux, certains nous étant même bénéfiques. Deux stratégies de survie : infecter ou persister L'évolution repose sur un principe fondamental : la survie du plus apte, autrement dit sur la compétition. Au jeu de la vie, dans un milieu où la concurrence est forte, un individu ne peut survivre que s'il s'adapte en permanence aux changements. S'il ne s'adapte pas, il est condamné. Vouloir survivre est un défi ambitieux : tous les organismes, y compris les virus doivent se maintenir dans le temps, répondre aux sollicitudes de l'environnement et notamment éliminer, échapper ou vaincre leurs ennemis et concurrents.
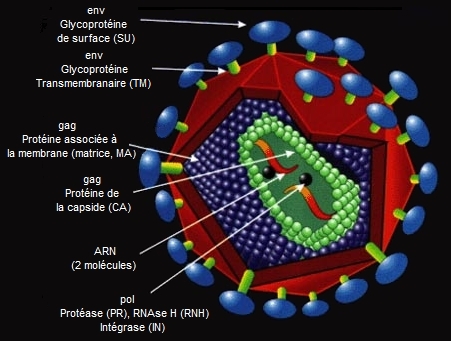
Dans le cas d'un virus, son ennemi est le système immunitaire de son hôte. Nous disposons de cellules sentinelles qui vont repérer les virus ou les cellules infectées par des virus. Ces sentinelles vont déclencher la production de lymphocytes (globules blancs). Soit la cellule a détecté la présence de virus et les lymphocytes T activent les lymphocytes B. Ceux-ci se transforment en plasmocytes producteurs d'anticorps qui vont neutraliser le virus. Soit la cellule est infectée et les lymphocytes T induisent la production de lymphocytes tueurs qui vont détruire la cellule infectée. Connaissant ce risque, si le virus ou plus généralement le parasite génétique veut survivre et maximiser ses chances d'avoir une descendance, deux stratégies s'offrent à lui : soit il agit seul mais doit compter sur un nombre important d'individus pour se multiplier à un taux élevé, soit il développe une stratégie de "persistance" consistant à être assimilé par son hôte tout en conservant ses fonctions génétiques. Le premier cas concerne le virus infectieux. Egoïste, il cherche à assurer sa propre survie. Il n'est pas attaché à un phénotype particulier et n'a donc aucun impact sur la compétition ou l'évolution de son hôte. De plus, l'avenir de son espèce n'est pas garanti (sauf mutation) car son action peut soit être contrée par le système immunitaire soit, si elle trop parfaite, conduire à l'extinction de l'espèce qu'il a pris pour cible. Ce type de virus ne nous intéresse pas car sa finalité est purement destructrice. Etre persistant pour survivre Le virus génétiquement assimilé par son hôte est plus intéressant. Pour survivre il va développer une stratégie consistant à s'incorporer dans un phénotype où il sera persistant. Les expressions "la survie du plus apte" ou "que le meilleur gagne" prend ici tout son sens. Pour le virus, les avantages sont multiples. En général le parasite génétique va co-évoluer en même temps que l'espèce qu'il a prise pour hôte. A l'image des parasites vivant en symbiose avec leur hôte, le virus persistant va également tirer profit des ressources de son hôte, sa survie étant assurée au prix d'un minimum de dépenses d'énergie qu'il va concentrer dans son champ d'activités, et enfin il va abandonner les tâches qu'il ne peut effectuer à son hôte. Ceci fait toute la différence entre les concepts d'égoïsme et de persistance. Le parasite génétique égoïste ne vise que son intérêt personnel, le parasite génétique persistant, au contraire, doit ajouter au génome de son hôte une nouvelle identité génétique moléculaire afin d'assurer sa pérénité tout en empêchant la compétition et tout déplacement de séquences. Mais cette compétition peut provenir du même parasite génétique que l'agent persistant lui-même. C'est pourquoi les virus génétiques persistants empêchent généralement leur propre réplication ou produisent des enzymes de restriction-modification (cas des phages P1 et P7) notamment pendant la réplication, forçant la cellule colonisée à maintenir le virus en bon état afin d'empêcher l'autodestruction de son hôte. A l'image de la symbiose, une fois de plus c'est le parasite colonisateur qui trouve un intérêt dans cette cohabitation forcée. Il peut s'installer durablement dans son hôte tout en continuant à lutter contre les autres agents parasites de manière plus statique. Quant à la cellule colonisée par un virus génétique, le virus a tout fait pour qu'elle y survive et continue à se reproduire, mais elle est à présent immunisée contre toute une série d'autres virus, y compris la famille des bactériophages T4[3].
Conséquence de cette stabilité, un virus génétique défectueux (dans lequel il manque un gène, incapable de se dupliquer par lui-même) est un vecteur de persistance idéal car il est souvent capable d'ajouter cet état de pérénité dans un parasite infectueux. Tirer profit des défectuosités génétiques représente donc une stratégie fréquemment utilisée par les virus pour persister durablement, dont l'effet ressemble fort au résultat que peut avoir un gène égoïste. Des virus créateurs de gènes Pour s'installer de manière durable dans son hôte, un virus doit être très compétitif et cette faculté nécessite la plupart du temps la présence de composants très spécifiques. Ainsi, le gène RRIA du T4 qui permet d'étudier l'aspect moléculaire le plus simple des gènes, ne présente aucune fonction spécifique dans son hôte, E.coli. Ce gène viral est uniquement présent afin de permettre au T4 d'infecter la cellule d'E.coli déjà occupée par un génome viral (le lysogène).
En fait, d'autres bactériophages porteurs du T-paire sont très conservateurs, contenant des gènes dont la fonction est inconnue et qui sont apparemment inutiles pour la réplication virale. En outre, ces gènes n'ont généralement pas d'équivalents dans les hôtes, suggérant qu'ils n'ont pas été "volés" du génome de ces organismes comme l'affirment les spécialistes des bactériophages. Ainsi, le bactériophage T4 contient 274 gènes dont 69 seulement sont essentiels au développement d'E.coli et dont 42 sont similaires à ceux de la base de nucléotides GenBank. La majorité de ces derniers gènes sont curieusement plus proches des gènes eucaryotes, y compris du groupe des introns (gène non codant) de type I découverts dans le gène T4 TK. Même constat chez le virus à ADN WSSV de la crevette grise. Il contient 184 gènes dont 11 seulement sont similaires à ceux de la base GenBank, la plupart intervenant dans l'expression des protéines de réplication de l'ADN. Plus étonnant, le phage archaea hyperthermophile AFV1 (Acidianus Filamentous Virus 1) et d'autres virus à ADN n'ont pratiquement pas de gènes, pas même de réplication, et ne correspondent à aucun élément de la base GenBank. Enfin, d'autres virus sont similaires dans leur degré d'innovation tout en n'ayant pas de similitudes avec les gènes de leur hôte, y compris la plupart des virus à ADN infectant les cellules eucaryotes. En fait, plusieurs lignages de virus présentent de nombreux gènes spécifiques à leur lignée. Ainsi, tous les représentants de la famille des baculovirus, des virus à ADN des insectes, présente un génome dont 80% des gènes sont uniques. On constate donc que la majorité des phages et des gènes viraux sont spécifiques à différentes familles de virus – et non pas vis-à-vis de leur hôte – bien que certaines similitudes frappantes existent chez les gènes eucaryotes. Ces virus persistants peuvent donc produire des effets non mortels pour leur hôte mais importants sur les plans génétique et, semble-t-il, de l'évolution. Cette acquisition du virus par son hôte s'effectue soit par recombinaison de brins d'ADN (ce qu'on appelle un transfert horizontal de gènes par transposons) soit par l'action d'un bactériophage suite à une infection lysogénique (le phage intègre son génome dans l'ADN de la cellule hôte). S'il est communément admis par les microbiologistes que les virus servent simplement de vecteur pour déplacer les gènes d'une cellule à l'autre, de plus en plus de virologues prétendent que les bactéries acquièrent la plupart de leur nouveaux gènes grâce aux virus de la cellule hôte. Mais une nouvelle découverte vient complexifier un peu plus toute cette théorie. Découverte d'obélisques dans le système digestif Des chercheurs de l'Université de Stanford ont découvert un nouveau type d'entité virale appelé "obélisque" dans le système digestif humain. Selon les auteurs, "Des séquences d'obélisques furent trouvées dans 7% des bactéries intestinales humaines et dans 50% des bactéries de la bouche" chez 472 personnes originaires principalement d'Amérique du Nord (cf. I.N. Zheludev et al., 2024; Nature, 2024).
Les obélisques présents dans les bactéries de différentes parties du corps ont des séquences distinctes. Leur génome présente apparemment des brins d'ARN de ~1 kb et des séquences propres potentiellement capables de fabriquer des protéines. Selon les auteurs, du fait qu'ils contiennent des gènes qui ne ressemblent à aucun de ceux découverts à ce jour dans d'autres organismes, "il s'agit d'une classe d'ARN variés qui ont colonisé et sont passés inaperçus dans les microbiomes humains." A ce jour, les spécialistes ne savent pas dans quelle mesure les obélisques pourraient affecter la santé humaine, mais ils pourraient altérer l'activité génétique de leurs hôtes bactériens, ce qui pourrait à son tour affecter les gènes humains. Pour les biologistes et virologues, une grande question est de savoir si les virus ont évolué à partir de viroïdes (les plus petits pathogènes viraux connus) et d'obélisques de plus en plus complexes, ou s'ils ont d'abord émergé puis dégénéré en ces structures plus simples. Selon la virologue Anamarija Butković de l'Institut Pasteur de Paris qui n'a pas participé à cette étude, les composants du microbiome "forment un équilibre. Il est intéressant de réfléchir à ce que font les obélisques et à la manière dont ils pourraient affecter tout cet équilibre." La réponse pourrait venir de la bactérie Streptococcus sanguinis qui réside dans la bouche humaine dans laquelle les auteurs ont découvert des obélisques. S.sanguinis étant facile à mettre en culture, Buković et ses collègues proposent que les scientifiques pourraient utiliser ces bactéries pour mieux comprendre comment les obélisques se répliquent, comment ils affectent les bactéries et ce que font leurs protéines. De telles expériences pourraient conduire à des découvertes sur l'origine de la vie elle-même. Certains scientifiques ont émis l'hypothèse qu'étant donné que les viroïdes et leurs proches parents sont petits, simples et ont la capacité de s'auto-répliquer, ils sont les précurseurs de la vie sur Terre. Même à présent que les scientifiques ont découvert les obélisques, ceux-ci y ont peut-être participé également voire façonné la vie dès le début. L'origine des cellules eucaryotes En tenant compte du taux relativement élevé de variations virales, de recombinaisons et d'adaptations (mutations), des virologues affirment que les virus plutôt que les bactéries sont véritablement capables d'explorer toute l'étendue des séquences d'ADN. Ils concluent que les virus sont les ultimes créateurs génétiques, inventant de nouveaux gènes en grands nombres, certains trouvant leur chemin dans des lignages d'hôtes grâce à une colonisation virale stable. Voyons cette théorie en détail. Si les virus et les bactéries ont évolué grâce à des mécanismes d'infections, où pourrait-on trouver des mécanismes similaires dans les cellules eucaryotes ? Aurions-nous négligé des adaptations génétiques par infection ou par transduction[4] ? Des agents parasites génétiques persistants pourraient-ils être impliqués d'une quelconque manière dans l'évolution eucaryotique ? Ou faut-il conclure que les structures complexes et les processus physiologiques sont apparus dans les cellules eucaryotes uniquement par les mutations ponctuelles, les échanges sexuels et la recombinaison ? Nous allons tenter de répondre à toutes ces questions. De nombreux organismes métazoaires portent une charge d'éléments génétiques parasites, le fameux ADN de rebut, dont on suppose qu'il s'est accumulé de manière régulière au cours de l'évolution. En comparant l'ADN entre espèces de différents règnes, et notamment celui des procaryotes avec les eucaryotes, on observe une augmentation étonnement faible du nombre de gènes qui passe par exemple de 2350 gènes chez E.coli à 8800 gènes chez la mouche pour atteindre environ 40000 gènes chez l'homme. On constate également que la densité de gènes chute de 90% de séquences codantes dans le génome d'E.coli ou de la mouche à moins de 3% chez l'homme.
L'excédant soit plus de 97% des gènes humains est constitué de séquences non codantes, répétées, d'éléments parasites, y compris de transposons de type I et II, ainsi que d'autres éléments parasites distinctifs tels que les LINEs et SINEs (Long et Short Interspersed Elements) qui sont clairement des traces de rétrovirus endogènes humains HERV (respectivement des séquences du gène pol et des séquences des régions LTR et pol). Le génome humain et celui d'autres vertébrés contient également plusieurs rétrovirus endogènes (les EVR) apparemment intacts et stables[5]. Ainsi, chez l'homme, le chromosome 21 porte 225 gènes codant pour des protéines, mais il porte également 2000 éléments rétroviraux endogènes ! Mais ce n'est pas exceptionnel en soi car la propagation des rétrovirus repose justement sur l'intégration de leur génome dans l'ADN humain pour se répliquer. Au total, l'ADN humain contient environ 8% de séquences d'anciens virus. En 2021, des chercheurs ont découvert que des séquences du SARS-CoV-2, le trop fameux Covid-19, avaient été intégrées par transcription inverse dans le chromosome X humain de patients guéris depuis longtemps de la Covid-19 mais apparemment recontaminés.
Au cours de la cartographie du génome humain, les chercheurs ont identifié 113 exemples de gènes humains inexistants chez les organismes eucaryotes plus simples alors qu'ils sont présents dans certaines espèces de bactéries. Mais il s'agit d'une observation logique si on considère qu'il y eut des transferts horizontaux directs de gènes entre les bactéries et les vertébrés. La plus grande discontinuité dans l'évolution entre les procaryotes et les eucaryotes apparaît dans les différences entre leurs protéines de réplication respectives. Les bactéries comme les eucaryotes utilisent un ensemble spécialisé de protéines pour répliquer leur ADN. Ces ensembles complexes de protéines très réactives effectuent des fonctions très similaires dans les deux règnes, alors qu'elles ont peu si non aucune séquence en commun et ne présentent aucun indice de forme dérivée d'un ancêtre commun.
Enfin, bien que les protéines de réplication des eucaryotes partagent quelques éléments avec leurs cousines comme certaines protéines de réplication archéobactériennes, ces deux ensembles diffèrent également de façon très marquée en ce qui concerne les complexes de reconnaissances de l'origine de l'ADN et les protéines des domaines de liaison à l'ADN non apparié (liant les simples brins ou monocaténaires). Prochain chapitre Similitudes entre certains bactériophages et eucaryotes
|
||||||||||||||||||||||||||||||||||||