|
|
|
Le rôle des virus dans l'évolution
Similitudes entre certains bactériophages et eucaryotes (II) Si les organismes procaryotes et eucaryotes présentent des différences fondamentales, il existe plusieurs similitudes frappantes entre certaines bactériophages et gènes eucaryotes. Ainsi, le gène polymérase de l'ADN du bactériophage T4 ressemble très fort à celui des eucaryotes. Celui du T4 est membre de la famille "Pol beta", sensible aux mêmes inhibiteurs que le "Pol delta" eucaryote et à la polymérase alpha (primase), ce qui a conduit Margarita Salsa et Luis Blanco de l'Universidad Autonoma de Madrid à suggérer que ces deux gènes avaient un ancêtre commun. T4 est un phage de terminaison (terminal, à l'image du codon stop), ressemblant beaucoup à ceux que l'on retrouve dans les archéobactéries (par ex. le phage H) et les cyanobactéries. Bien que le T4 infecte E.coli et soit donc lytique, d'autres virus proches peuvent également infecter certaines espèces d'algues. Parmi ces virus, les Feldmania sont persistants mais l'infection est non apparente tandis que les Chlorella et les CSV-1 présentent dans les microalgues sont infectieux. Les espèces Chlorella sont des unicellulaires parasites, haploïdes[6] et asexués[7]. CSV-1 est un virus contenant un double brin d'ADN (380 kbp), dont les gènes codent une multitude d'enzymes (polymérase, kinase, synthase, dismutase, reductase, transposase, etc) à différentes étapes de la reproduction, y compris des enzymes de restriction et de modification qui sont rares chez les eucaryotes. Son génome contient également 80 introns ou gènes non codants. La jonction évolutionnaire entre les microalgues et les algues brunes filamenteuses coïncide également avec la bifurcation qui est apparue après le rayonnement du Précambrien (qui se termina voici 542 millions d'années), lorsque les métazoaires et la reproduction sexuée ont émergé. Analyse de l'ADN polymérase Les virus à ADN persistants des algues sont-ils à l'origine des protéines de réplication eucaryotes, reliant les différents embranchements de l'arbre universel de la vie ?
Pour examiner cette possibilité, il faut analyser les séquences de protéines et établir des dendogrammes (la classification arborescente) et cela devient très technique. Luis P. Villarreal, directeur du Center for Virus Research à l'Université de Californie à Irvine étudie le rôle des parasites génétiques tels que les virus dans l'évolution des organismes. Avec son équipe, il a étudié l'ADN polymérase (Pol), une enzyme impliquée dans le processus de réplication (type I) et de réparation (type II) de l'ADN. Villarreal a utilisé le programme TBLASTN[8] afin de comparer la séquence Pol du virus Feldmania, spécifique à cette espèce et présente dans les algues brunes filamenteuses avec une série d'autres Pol, y compris celles de quatre autres familles de virus à ADN (phycodnavirus, herpesvirus, poxvirus, baculovirus, avec sous-clades alpha, beta et gamma), de plusieurs bactériophages et avec deux ensembles distincts d'ADN archéobactériens Pol II (des ADN polymérases procaryotes impliquées dans la réparation de l'ADN). Il ressort de cette analyse que pratiquement chaque membre de l'ADN polymérase de la famille B est similaire et a conservé des domaines fonctionnels[9] au sein de toutes ces protéines. En outre l'analyse statistique des dendogrammes de ces protéines met en évidence des ensembles identiques ou clades de polymérases apparentées[10]. La plupart de ces clades présentent des distinctions et viennent se placer au centre (non résolu) de l'arbre de vie, à une exception près. L'ADN Pol du virus Feldmania est situé à la base du clade correspondant à celui de la cellule hôte de l'ADN Pol delta. Etant donné que ces arborescences n'établissent pas de polarité des caractères, ce résultat suggère que : - l'ADN Pol du virus Feldmania représente le géniteur de toutes les cellules ADN Pol delta ou, - le virus Feldmania a acquit sa version de l'ADN Pol d'un hôte qui ressemble au géniteur. Un géniteur d'origine virale semble le plus probable. Tous les autres ADN viraux Pol analysés ne font pas partie du clade correspondant à leur hôte bien que sur le dendogramme ces virus soient aussi âgés que Feldmania et soient apparentés aux bactériophages. En outre, si le virus Feldmania a hérité ou plutôt reçu son Pol de son hôte, ce serait inhabituel comparé au fonctionnement d'autres virus à ADN. Etant donné que ces virus sont transmissibles, il semble plus simple d'imaginer que le transfert s'est produit du virus vers la cellule hôte. De plus, suggérer qu'un virus fut à l'origine des protéines de réplication eucaryotes aiderait à comprendre la discontinuité entre les protéines de réplication bactériennes et eucaryotes, et pourrait également expliquer comment les eucaryotes sont reliées au monde bactérien. Notons que Patrick Forterre de l'Université de Paris-Sud à Orsay a émis des idées similaires. Analysons à présent d'autres gènes de phycoadnvirus, en particulier la dismutase superoxyde (SOD) et PCNA. Bien que ces résultats soient moins convaincants du fait que les données sont moins nombreuses, elles renforcent la thèse d'une origine virale de ces gènes.
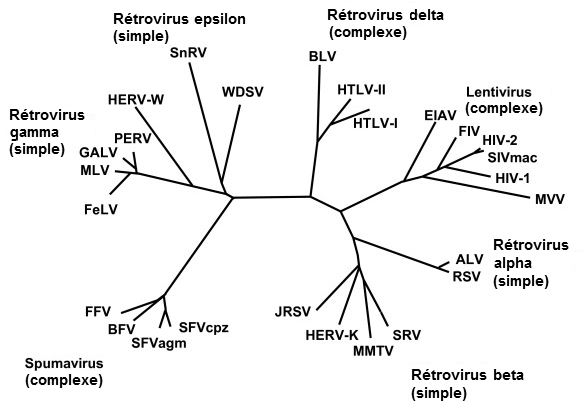
Des rétrovirus dans le placenta des mammifères Particularité des animaux placentaires Les mammifères peuvent également utiliser les rétrovirus endogènes pour faciliter le développement de l'embryon et supprimer l'activité du système immunitaire in utero. Comment les mammifères peuvent-ils tolérer cette présence étrangère auprès d'un embryon ? Pour comprendre ce processus complexe il faut d'abord définir le phénotype des mammifères et comprendre comment se forme le placenta. Tous les génomes mammaliens présentent des ensembles spécifiques et distincts de rétrovirus endogènes (ERV) et un très grand nombre de dérivés de rétrovirus défectueux, suggérant que les génomes mammaliens furent colonisés par des lignées spécifiques d'ERV longtemps avant que les espèces placentaires se séparent les unes des autres. D'après les analyses phylogénétiques, ces gènes rétroviraux furent introduits chez les primates voici quelque 40 millions d'années tandis que des gènes homologues trouvés chez les rongeurs et notamment chez la souris sont présents depuis 20 millions d'années. Les exemples comprennent les rétrotransposons (des séquences d'ADN endogènes transposables) LINE-1, SINE R, de nombreux transposons Alu, les apparentés aux HERV-K, l'IAP de souris, l'IAP du hamster, le rétrovirus Rd114 félin ainsi que le rétrovirus rhesus-Mason-Pfizer. La nomenclature des rétroposons et des ERV est plus confuse. Le projet HUGO indique qu'il existe plusieurs centaines de milliers d'ERV chez l'homme et qu'ils sont répartis en 24 familles. Nous avons à la fois des anciennes souches comme ERV-L et des versions récemment acquises d'ERV, telles les huit membres d'ERV K qui distinguent les humains des primates les plus proches. Toutefois, les espèces aviaires ne présentent pas cette importante colonisation de rétroposons, ce que confirme David Mindell de l'Université de Michigan à Ann Harbor. De plus, les mammifères correspondent génétiquement à leurs ERV alors que ce n'est pas le cas chez les oiseaux. Les mammifères et les oiseaux se différencient également par le fait que bien que leur embryons respectifs puissent subir l'infection génomique de rétrovirus, les mammifères peuvent inhiber l'expression de leurs gènes (par le biais de la méthylation de l'ADN) dans les tissus embryonnaires. Bien que des versions dérivées et défectueuses de ces ERV, en particulier celles où le gène env (Enveloppe) est supprimé, ne codent généralement aucun produit, l'analyse des codons synonymes[11] indique qu'un beaucoup plus petit nombre d'ERV intacts subsistent avec tout leur potentiel de codage. Rôle du placenta Tous les mammifères développent un placenta dérivé du trophectoderme (cellules externes de la morula) qui permet au blastocyste (œuf fécondé) de progresser et de s'implanter (nidifier) dans la paroi de la cavité utérine. Le placenta est constitué par les replis de la membrane (villosités) entourant le fœtus et de la muqueuse utérine.
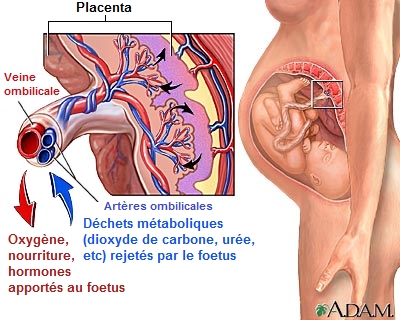
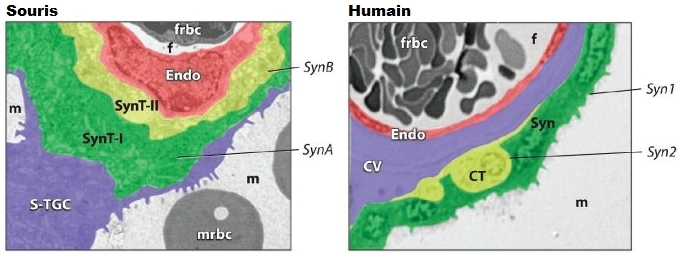
Le placenta est un organe en forme de tarte très vascularisé qui établit un contact sanguin avec la mère à travers la paroi utérine. Au début de la grossesse (la gestation chez les autres animaux) cet amas de chair très spongieux à travers lequel transite le sang remplace une partie des organes du foetus en formation. Le placenta apporte à l'embryon tout ce dont il a besoin (oxygène, eau, nutriments) et se charge d'éliminer le gaz carbonique et les déchets (urée, etc). Il contrôle également les niveaux hormonaux durant la grossesse et protége l'embryon contre toute réponse innée et immunitaire adaptative de la mère. Le placenta un organe foetal et non un organe maternel dont la durée de vie est limitée à environ 9 mois. Du point du vue immunologique, le placenta est donc un corps étranger que la mère tolère pendant la grossesse. C'est ici que l'on découvre le rôle inattendu des rétrovirus. Nous avons expliqué que les rétrovirus ont la faculté de fusionner la membrane de leur enveloppe avec l'enveloppe cellulaire de leur hôte. Cette fusion de deux membranes est facilité par l'action des syncytines qui interagissent avec la surface de la cellule "infectée". Ce mécanisme de fusion membranaire permet de former des structures géantes appelées "syncytia" constituées par la réunion de cellules individuelles. Or, le placenta présente justement cette structure à l'interface entre le foetus et la mère. Localisation et rôle des syncytines Parmi les protéines importantes codées par le gène env durant la grossesse, il y a la syncytine, une protéine associée aux HERV très active au moment de la nidation car elle est indispensable à la formation du placenta. Le trophecdoderme est la première cellule qui se différencie du blastocyste (l'oeuf fécondé) au bout de 3.5 jours, lorsque les cellules totipotentes de la masse interne (cellules souches embryonnaires) sont déterminées. Dans les années 1970, les chercheurs ont constaté que chez l'être humain normal et d'autres mammifères le trophectoderme ou les cellules placentaires produisaient des particules rétrovirales endogènes, des syncytines en grandes quantités. Plus tard, d'autres chercheurs découvrirent que la plupart des mammifères exprimaient des ERV correspondants dans les tissus placentaires et embryonnaires. De plus, les séquences HERV exprimées ou transcrites par le foetus et dans le placenta sont variées, certaines étant intactes, d'autres défecteuses. Les tissus placentaires et de l'embryon sont de loin les sites les plus communs et les plus abondants de l'expression des ERV, suivis par les tissus lymphatiques et les tumeurs malignes. Eric Larsson et ses collègues de l'Université d'Uppsala en Suède ont observé l'expression de la protéine env de l'ERV-3, une protéine de type syncytine dans les cellules placentaires d'un humain normal et ont suggéré qu'un ERV-3 intact est nécessaire, probablement pour l'immunosuppression (la suppression des réactions immunitaires). En effet, en "désactivant" le système immunitaire localement et temporairement, le foetus peut se développer in utero sans susciter de rejet de la part de la mère. Villarreal généralise ce concept du fait de la large colonisation des espèces placentaires par des ERV intacts et la nature généralement immuno-suppressive de la région hydrophobe membrane-voisine (membrane-spanning) env de l'ERV.
Bien que la protéine env de l'ERV-3 ait subit une mutation chez 1% des Caucasiens, la complexité et la quantité d'autres membres de la famille ERV suggère qu'elle et d'autres protéines env des ERVs sont exprimées et pourraient être impliquées dans la suppression immunitaire et d'autres processus vitaux du développement. Le virologue Thierry Heidmann de l'Institut Gustave Rousssy (IGR) de l'Université de Paris-Sud 11 supporte également cette hypothèse. Selon ses études, ce mode particulier de développement des mammifères semble trouver son origine dans l'introduction d'un rétrovirus dans le génome d'un ancêtre commun. Cet évènement se serait produit il y a quelque 100 millions d'années, avant même l'apparition des gènes rétroviraux de syncytine. "Il est possible que des infections successives se soient produites au cours de l'évolution et que les gènes nécessaires à la placentation aient été progressivement remplacés au sein de chaque espèce", explique Thierry Heidmann. Cela expliquerait aussi pourquoi le mécanisme placentaire ne fonctionne pas strictement de la même manière chez tous les mammifères. Des rétrovirus protègent l'embryon L'idée que des ERV seraient quelque part impliqués dans la protection des embryons des mammifères est plausible et attrayante. Mais pour tester cette idée, il faudrait supprimer toutes les familles d'ERV des embryons. Quand on sait qu'il existe des milliers de loci[12] d'ERV, ce travail est impossible et donc utopique. Toutefois, les découvertes faites sur les polyomavirus à la fin des années 1970 nous fournissent un moyen de résoudre ce problème. En effet, ces études indiquent que lorsque des cellules cancéreuses embryonnaires (carcinome embryonnaire ou EC) expriment l'antigène T (T-Ag) du polyomavirus SV40 (Simian Virus 40), il bloque l'expression des rétrovirus endogènes sans affecter la différentiation de la cellule cancéreuse. De plus, T-Ag s'exprime dans les cellules souches embryonnaires sans affecter leur différenciation. Cependant, les lignées transgéniques ne parviennent pas à s'établir, suggérant que l'expression du T-Ag perturbe l'expression de l'ERV sans affecter la différenciation cellulaire.
Pour tester cette hypothèse, Villarreal et ses collègues ont modifié sélectivement l'expression des gènes des embryons de souris dans des cellules cancéreuses, bien que des cellules souches aient été préférables mais ne se différenciant pas suffisamment longtemps dans le trophectoderme. Ils ont évalué de quelle manière le T-Ag affectait l'expression de l'IAPE-A, un ERV qui exprime la protéine env. Nous savons que la séquence env s'exprime très fortement dans le blastocyste (stade 5-7 jours) d'une souris normale. Mais dans les cellules cancéreuses, T-Ag bloque l'expression de l'IAPE-A sans affecter la différenciation des EC dans les corps embryonnaires, qui ressemblent forts à des blastocystes de 3.5 jours. Bien que les corps embryonnaires dérivés des EC ne soient pas totalement fonctionnels et ne produisent pas de résultats viables (en raison du manque de cellules totipotentes), ils poursuivent le programme d'implantation alors que les corps embryonnaires exprimant T-Ag ne parviennent pas à s'implanter. Ces résultats renforcent l'idée que les ERV jouent un rôle durant l'implantation de l'embryon, leur permettant peut-être d'éviter d'être reconnu par le système immunitaire de la mère. Bien que ces résultats ne soient pas définitifs, cela peut signifier que le placenta des mammifères aurait évolué grâce à la colonisation de rétrovirus endogènes qui auraient apporté des phénotypes complexes dans les embryons, en particulier via leur placenta. Ainsi, comme le pensent Villarreal, Heidmann, Larsson et consorts les embryons placentaires se comportent apparemment comme des parasites qui envahissent et infectent leur hôte – leur mère – y trouvant leurs moyens de subsistance et produisant des virus locaux afin de supprimer les réponses immunitaires. Cette relation ressemble à un phénomène observé chez les guêpes qui implantent leurs œufs dans les larves de chenilles hôtes. Les défenses innées du hôte sont neutralisées par des virus génomiques endogènes (polyadnavirus) fabriqués au sein même des cellules nourricières de la guêpe entourant les œufs et bloquant les réponses de défense antiparasite de la chenille hôte. Ensuite, de ces larves infectées éclosent des guêpes volantes ordinaires. Aussi, il n'est pas exagéré de penser que de tels mécanismes parasitaires puissent être à l'origine ultime de la morphogénèse, si commune chez de nombreux insectes volants. Si nous voulons découvrir d'autres phénomènes de l'évolution dans lesquels des organismes ont acquis des phénotypes complexes et très adaptatifs, nous devons chercher des traces de parasites génétiques persistants. Après tout, les recombinases RAG1 et RAG2, essentielles à l'origine et au fonctionnement du système immunitaire adaptatif des mammifères ressemble fort à une intégrase rétrovirale. Le locus de la région chromosomique du complexe majeur d'histocompatibilité (MHC) est lui-même suspect quand on sait qu'il a été colonisé 10 fois plus fréquemment sur le plan génomique que les ERV[13].
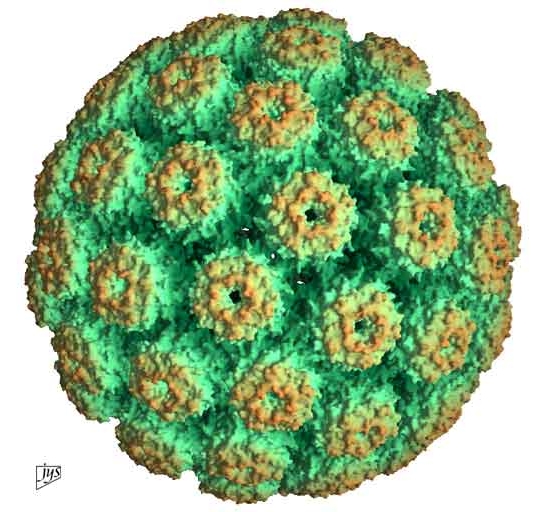
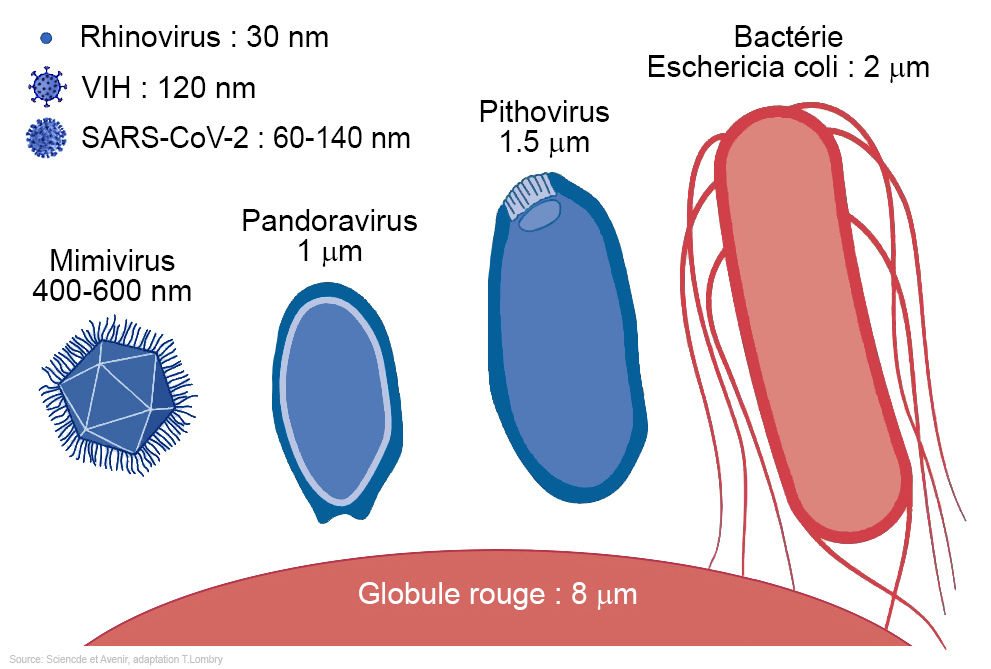
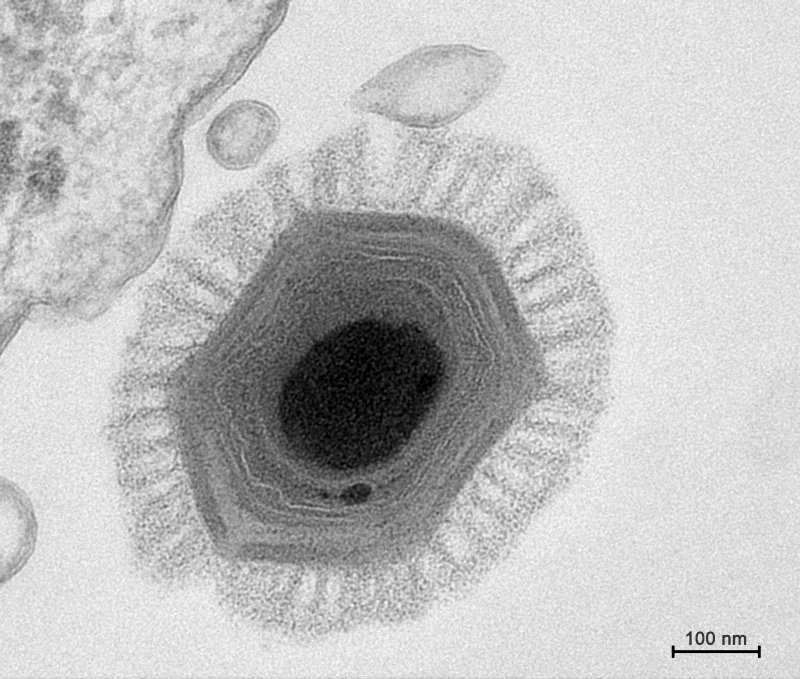
En résumé, pour survivre et ne pas tuer leur hôte, les rétrovirus ont dû s'adapter. Chaque rétrovirus a pénétré la cellule de son hôte et sectionné son ADN pour y placer sa séquence génétique. Au fil des générations cette mutation s'est multipliée et permis de créer le placenta qui protège l'embryon. Cette stratégie biologie donna un avantage aux mammifères placentaires sur ceux qui étaient ovipares notamment. Finalement, cette invention renforça le lien entre la mère et son bébé. Chez les mammifères, le bébé qui naît est presque prêt à conquérir le monde, ce qui lui donne un sérieux avantage sur tous les autres animaux non placentaires. En 1992, des chercheurs ont découvert un pandoravirus parasitant une amibe vivant dans l'eau d'une tour de refroidissement de Bradford, en Angleterre (cf. B.La Scola et al., 2003). Il mesurait 400 nm de diamètre et présentait une capside icosaédrique (à 20 faces); il s'agissait d'un virus géant proche du mimivirus. Depuis, de nombreux variants du mimivirus ont été identifié dans différentes régions du monde. Comme le rappellent Brandes et Linial dans un article publié dans la revue "Viruses" en 2019, "les virus géants sont des virus infectant les cellules eucaryotes et possédant au moins 500 gènes codant pour des protéines". Leur diamètre dépasse 200 nm et leur génome possède plus de 300000 paires de bases. On connaît actuellement 48 virus géants dont la majorité vit dans des milieux aquatiques. Parmi ceux-ci, en 2017 des chercheurs découvrirent 16 nouveaux virus géants (dont Dasovirus, Harvovirus, Hyperiovirus, Solumvirus et Sylvanvirus) par hasard dans un échantillon de sol prélevé dans la forêt d'Harvard aux Etats-Unis dans le cadre d'une étude sur le climat (cf. F.Schulz et al., 2020). Les chercheurs ont notamment découvert de nouvelles lignées virales ou proches d'autres virus géants comme les Klosneuvirus, les Tupanvirus et le virus CroV de la famille des Mimiviridae dont le plus connu est le mimivirus. Ces virus géants comprennent jusqu'à 240 gènes codant pour des protéines de la capside. A consulter : List of the main “giant” viruses, IGS/CNRS, 2019
Sachant que ces 16 nouveaux virus géants furent identifiés dans un seul échantillon de sol, les chercheurs estiment que ces micro-organismes sont bien plus nombreux et répandus sur Terre et se comptent probablement par milliers. C'est tout un écosystème microbien dont on sait encore peu de choses. Heureusement, sur base d'isolats viraux (des virus isolés de leur hôte), la microbiologie a permis de mieux comprendre la diversité et la biologie des virus géants. Leur culture en laboratoire a permis de mieux comprendre leur morphologie, leurs fonctions et leurs stratégies d'infection (cf. S.W. Wilhelm et al., 2017; F.Schultz et al., 2022). Le mimivirus ou "mimicking virus" (virus mimétique) présenté ci-dessous fut découvert en 2003 par une équipe du CNRS dans des amibes provenant des circuits de refroidissement d'eau de systèmes de climatisation (cf. D.Raoult et al., 2003; D.Raoult et al., 2004). Le mégavirus qui lui est apparenté fut découvert en 2011 dans du plancton au large du Chili, près de la station biologique marine de La Cruces (cf. J.-M. Claverie et al., 2011).
Ces deux virus géants possèdent un double brin linéaire d'ADN, ils ont 594 gènes en commun - par comparaison, le virus de la grippe ne possède qu'une dizaine de gènes - et possèdent des enzymes pour traduire l'ADN et fabriquer des protéines, ce que ne possèdent pas les virus traditionnels qui doivent compter sur les enzymes des cellules hôtes qu'ils infectent pour les fabriquer. Ainsi le Megavirus chilensis est capable de fabriquer 1120 protéines. Nous verrons que plus récemment, des chercheurs ont découvert des virus géants dans le permafrost de Sibérie qui étaient congelés depuis plus de 30000 ans et sont parvenus à les réanimer. C'est le cas du Pithovirus et du Mollivirus. Mais comment ces virus géants ont-ils hérités de ces protéines et de ces enzymes ? D'un lointain ancêtre où d'une cellule hôte ? L'existence de ces virus géants chez les animaux et les plantes renforce l'idée que les virus se seraient développés très tôt dans l'évolution et que certains nous aident à nous développer et nous protéger des infections. Certains, car en 2005 on rapporta un cas de pneumonie chez l'homme provoqué par le mimivirus qui provoque également la même maladie chez des souris (cf. D.Raoult et al., 2005).
Le mimivirus donne l'impression d'imiter les bactéries car il est plus gros que beaucoup d'entre elles (plus de 1.2 million de paires de bases et 400 nm de diamètre). Outre cette particularité, il possède 7 gènes communs aux domaines (anc. règnes) des bactéries, des archées et des protistes (eucaryotes). Le mimivirus apparaît sur les dendrogrammes sur une quatrième branche proche de l'origine des eucaryotes mais distincte des autres domaines. Ceci suggère une très grande ancienneté. Il est donc possible de réaliser un arbre phylogénique des êtres vivants incluant le mimivirus et des dizaines d'autres mégavirus et donc l'ensemble des virus. Selon les dernières études, le génome humain comprend 2% de gènes eucaryotes, 8% de rétrovirus, ceux-là même qui nous aident à nous protéger des infections et 43% de gènes d'origine virale ! Dernier chapitre Arc où le témoin de l'origine virale de notre mémoire
|
||||||||||||||||||||||||||||||||||||||||||||||||||||