|
|
|
L'origine et l'avenir de l'Homme
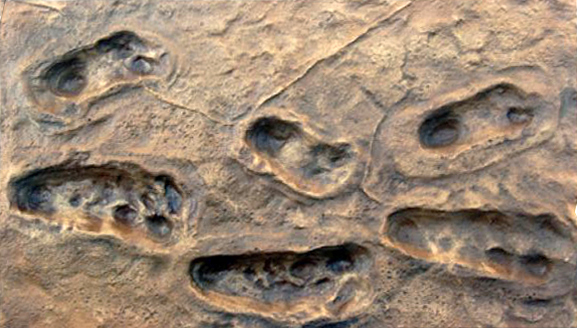
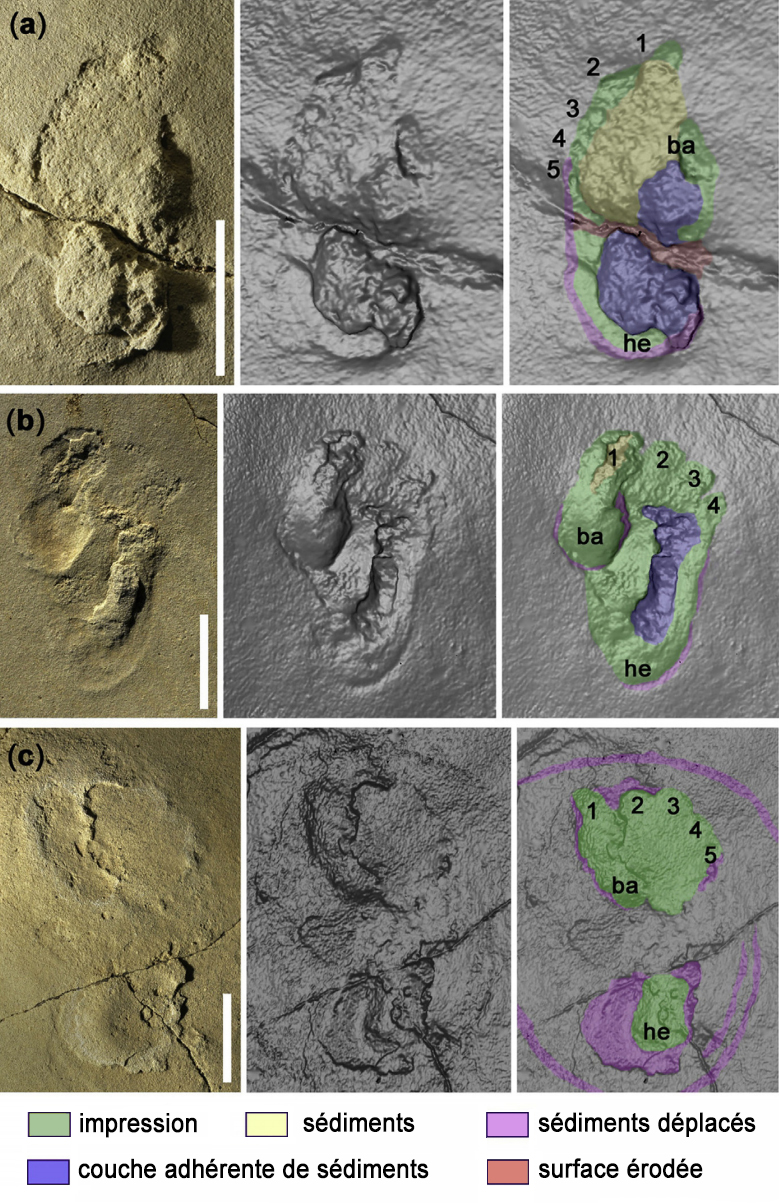
L'origine de la bipédie (V) 1. La piste de Laetoli En 1978, Mary Leakey découvrit des empreintes de pas à 45 km au sud des gorges d'Olduvai, en Tanzanie (cf. Google Map). Elle furent datées de 3.7 millions d'années. Elles appartiendraient à des homininés de l'espèce Australopithecus afarensis. Comme on le voit sur les photographies suivantes, les longueurs des empreintes mesurent entre 18.5 et 21.5 cm pour une foulée de respectivement 28.7 et 47.2 cm. Elles représentent les pas d'un adulte mesurant entre 1.34-1.56 m accompagné d'un enfant mesurant entre 1.15-1.34 m ainsi que celles d'un petit animal qui suivit une autre direction mais le site comprend au total les empreintes d'une vingtaine d'espèces d'animaux (hyènes, félins, babouins, girafes, gazelles, rhinocéros, antilopes, lièvres, oiseaux, etc). La conservation de telles empreintes est exceptionnelle et tient d'une série extraordinaires de hasards. Le volcan de Sadiman visible à 20 km de distance est entré en éruption quelque temps avant le passage des préhumains et éjecta beaucoup de cendres. La pluie s'abattit ensuite sur la région (des traces de gouttes de pluie furent mises à jour). Ces hominidés ont marché dans ces cendres volcaniques humides qui conserva leurs empreintes aussi précisément que du plâtre. Une seconde éruption volcanique recouvrit leurs traces sous 30 mètres de cendres, les protégeant ainsi de l'érosion. Au fil du temps, à l'abri de l'oxydation et de l'humidité, les empreintes finirent par se minéraliser. Ensuite, au gré des aléas météorologiques, la couche fut à nouveau exposée à l'érosion ce qui permit de découvrir les empreintes à seulement quelques dizaines de centimètres sous la couche de terre. En 1979, divers moulages dont un en silicone furent réalisés par les chercheurs du Musée National de Tanzanie et du Metropolitan Museum of Arts de New York puis les empreintes furent recouvertes pour les préserver de l'érosion - de la Mousson notamment - et des pilleurs. La "piste de Laetoli" fut réexaminée entre 1992 et 1996. Lors de la première mission, une bande de 3x3 mètres fut étudiée et un nouveau moulage aussi précis que possible fut réalisé. En 1993, on découvrit que des racines d'arbres risquaient de détruire les empreintes. Pour les besoins de la Science et des générations futures, on décida de laisser les traces fossilisées sur place et d'empoisonner les arbres pour préserver les empreintes. Depuis 1996, l'emplacement ressemble à une zone herbeuse rendue à l'état sauvage.
Grâce à ces empreintes, on sait donc avec certitude que les préhumains vivant il y a environ 3.7 millions d'années étaient bipèdes et que cette démarche ne posait aucun problème aux enfants. On peut donc supposer qu'ils ont hérité de cette faculté quelques centaines de milliers d'années plus tôt sinon davantage. 2. La piste crétoise de Trachilos En 2002, alors qu'il était en vacances en Crète, le paléontologue Gerard Gierlinski de l'Institut géologique polonais découvrit sur le rivage rocheux de Trachilos situé au nord-ouest de la Crète, des empreintes fossilisées dans des sédiments que l'on voit ci-dessous. Elles furent datées de la fin du Miocène, il y a environ 5.7 millions d'années. A cette époque l'endroit étant un rivage sablonneux, peut-être le petit delta d'une rivière. Dans une étude publiée par Gierlinski et ses collègues en 2010, les chercheurs sont arrivés à la conclusion qu'il s'agit d'empreintes d'homininés plutôt que d'hominidés et qu'il ne s'agissait pas non plus d'empreintes de singes (Pan) en raison de la disposition du gros orteil. La date fut confirmée dans une nouvelle étude publiée en 2017 par la même équipe dans les "Proceedings of the Geologists' Association".
Cette découverte soulève une nouvelle énigme : est-il possible qu'il existait déjà des préhumains dans le sud de l'Europe 2 millions d'années avant Laetoli ? Et de quelle espèce préhumaine peut-il bien s'agir sachant que les plus anciennes, Sahelanthropus tchadensis alias Toumaï (7 Ma) vivait au Tchad et Orrorin tugenensis (6 Ma) vivait en Afrique de l'Est ? De plus, à la fin du Miocène, la plupart des espèces d'homininés étaient assez proches du genre Pan, ce qui ne semble pas être le cas de ces nouvelles empreintes dont les orteils sont tous alignés sans le moins écart. Précisons que les pieds humains ont une forme très distincte, différente de celle de tous les autres animaux terrestres. La combinaison de longs métatarses, d'une longue voûte plantaire, de cinq orteils courts (tarses) sans griffes et d'un hallux (gros orteil) plus grand que les autres orteils est unique. Les pieds de nos ancêtres les plus proches, les grands singes, ressemblent davantage à une main humaine avec un hallux en forme de pouce qui se détache sur le côté et est opposable aux autres orteils pour assurer une parfaite préhension des objets. Les traces des présumés A.afarensis de Laetoli sont assez semblables à celles des humains modernes, sauf que le talon est plus étroit et l'empreinte ne présente pas de voûte plantaire. En revanche, Ardipithecus ramidus âgé de 4.4 millions découvert en Éthiopie présente un pied semblable à celui du singe. Les recherches ont conduit les spécialistes à conclure qu'Ardipithecus est un ancêtre direct des homininés ultérieures, ce qui implique que le pied humain n'avait pas encore évolué à cette époque là.
Pourtant les empreintes de Trachilos sont 1.3 million d'années plus anciennes que celles d'A.ramidus et ont une forme indubitablement humaine. C'est particulièrement vrai pour les orteils. Le gros orteil est semblable à celui de notre propre espèce, en taille comme en position; il est également associé à une boule plantaire qui n'est jamais présente chez les singes. La plante du pied est proportionnellement plus courte que dans les traces de Laetoli, mais elle a la même forme générale. En bref, la forme des empreintes de Trachilos indique sans ambiguïté qu'elles appartiennent à un homininé précoce, un peu plus primitif que celui de Laetoli. Selon Per Ahlberg de l'Université d'Uppsala qui est coauteur de l'article publié en 2017, le problème que posent ces empreintes est leur âge et l'endroit de leur découverte. Si elles sont plus jeunes que Toumaï et contemporaines d'Orrorin mais plus anciennes que A.ramidus, c'est en contradiction avec l'hypothèse selon laquelle A.ramidus est un ancêtre direct des homininés ultérieurs. En outre, jusqu'en 2017, tous les homininés fossiles âgés de plus de 1.8 million d'années (l'âge des premiers fossiles d'Homo georgicus découvert en Géorgie) proviennent d'Afrique, ce qui conduit la plupart des chercheurs à conclure que c'est en Afrique que le groupe a évolué. Cependant, les traces de Trachilos sont correctement datées par une combinaison de foraminifères (microfossiles marins) présents dans les lits supérieurs et sous-jacents, plus le fait qu'elles se situent juste en dessous d'une roche sédimentaire très distincte formée lorsque la mer Méditerranée s'est brièvement asséchée il y a 5.6 millions d'années. Par une curieuse coïncidence, comme nous l'avons évoqué, début 2017 une autre équipe de chercheurs avait conclu que les premiers fragments de Graecopithecus découverts en Grèce et en Bulgarie appartenaient à un homininé âgé de 7.2 millions d'années. A l'époque où les traces de Trachilos se sont imprimées dans les sédiments, à la fin du Miocène, le désert du Sahara n'existait pas; la région ressemblait à une savane qui s'étendait de l'Afrique du Nord vers l'est de la Méditerranée orientale. En outre, la Crète ne s'était pas encore détachée de la péninsule grecque. Il est donc facile d'imaginer comment les premiers homininés ont pu progresser dans toute l'Europe du Sud-Est et en Afrique, laissant au passage leurs traces sur un rivage méditerranéen qui allait former une partie de l'île de Crète. En conclusion, comme le dit Ahlberg "cette découverte contredit l'histoire établie de l'évolution humaine primordiale et va probablement susciter de nombreux débats". Mais la communauté des paléoanthropologues n'est peut-être pas prête à reviser son histoire sans autre preuve de la présence d'homininés en Crète au Miocène. Ces découvertes ainsi que l'analyse morphologique des différentes espèces préhumaines, et en particulier de Toumaï, Orrorin et des Australopithèques, nous conduisent à nous demander pourquoi nos ancêtres se sont-ils redressés et sont-ils descendus des arbres ? La réponse fait encore l'objet de nombreux débats. A défaut d'une explication définitive, nous en sommes réduits à des conjectures et des analyses comparées dont voici un résumé. La convergence génétique Il existe quatre mouvements associés à la bipédie : la station debout, la marche, la course et le saut ou le bond. Les déplacements sur deux pieds (ou deux pattes) est une capacité qui est apparue à de multiples reprises au cours de l'évolution. Certaines espèces sont bipèdes par obligation (elles n'ont pas d'autre alternative) d'autres le sont facultativement, dans certaines circonstances. Parmi les animaux bipèdes citons certains espèces d'archosaures, de dinosaures, les oiseaux, les macropodes comme l'autruche, le rat kangourou et bien sûr la plupart des hominidés. Certains lézards se redressent également sur leurs pâtes arrière pour s'enfuir plus rapidement, de même que certains petits mammifères comme les suricates lorsqu'ils surveillent leur territoire mais ils se remettent à quatre pattes pour courir. Chez nos ancêtres, la bipédie remontant à plus de 6 ou 7 millions d'années, elle n'est pas conditionnée par le développement du grand cerveau des espèces Homo ni par l'invention des outils de pierre (premiers galets et bifaces taillés) au Paléolithique. Etant donné que la bipédie est présente chez d'autres animaux mais uniquement chez les vertébrés, il s'agit d'une convergence génétique spécifique probablement apparue suite une ou des influences extérieures comme la pression environnementale, mais pas uniquement. Explications. La pression environnementale : la "théorie de la Savane" De quelle pression environnementale peut-il s'agir et fut-elle unique ? Aujourd'hui il est encore très difficile de répondre à ces questions. La "théorie de la Savane" généralement évoquée trouve son origine dans l'interprétation des études du paléosol et du paléoclimat de l'Afrique orientale. L'étude géologique nous apprend que la formation du Rift africain débuta au Miocène, il y a environ 23 millions d'années (cf. la charte stratigraphique) et modifia notamment le climat dans l'Est de l'Afrique. Thure E. Cerling, géologue à l'Université d'Utah et ses collègues ont publié en 2011 dans la revue "Nature" une très intéressante étude sur l'évolution du sol et indirectement du climat en Afrique orientale à l'époque des homininés, grosso-modo entre deux millions d'années avant A.ramidus jusqu'à l'apparition des Homo erectus et Homo sapiens.
Comme nous l'avons évoqué à propos du changement climatique survenu à l'époque de Graecopithecus, comme on le voit dans les graphiques ci-dessus résumant les analyses des chercheurs, ils ont découvert qu'il y a 7 millions d'années, la partie orientale de l'Afrique allant du bassin de l'Omo-Turkana au Kenya et en Ethiopie à la vallée d'Awash en Ethiopie, où les fossiles des premiers préhumains furent découverts, était couverte de prairies ponctuées ci et là de quelques arbres épars. C'était une savane arborée comme on en trouve aujourd'hui au Kenya ou en Tanzanie par exemple. La couverture boisée s'est accentuée au cours des quelques millions d'années qui suivirent, atteignant sa plus grand extension il y a environ 3.6 millions d'années, époque à laquelle les bois occupaient entre 40 et 60% de la région étudiée. Ensuite ces bois se sont réduits. Vers 1.9 million d'années, en aucun endroit la superficie des bois ne dépassa la moitié du territoire. Le paysage continua ensuite à s'ouvrir, une tendance qui perdure encore aujourd'hui. On présume que ce changement de climat et l'ouverture des forêts auraient poussé les premiers hominidés à descendre des arbres pour explorer la savane et y chasser les animaux. Si cette théorie est valable pour l'Afrique orientale, elle n'explique pas à elle seule la bipédie puisque A.ramidus vivait probablement dans la forêt, Orrorin était arboricole et devait probablement apprécier les arbres tandis que Toumaï découvert à plusieurs milliers de kilomètres à l'ouest du Rift vivait dans une région lacustre qui s'est progressivement désertifiée. Si ces trois espèces avaient une prédisposition génétique pour se redresser et marcher, l'environnement et les climats respectifs différents des deux régions impliquent une cause sans rapport direct avec le climat. Nos ancêtres ont eu la chance de se séparer des autres hominidés vivant dans la forêt. Aujourd'hui, tous les grands singes, chimpanzés, bonobos et gorilles vivent dans la forêt. Ils ne font jamais d'excursions dans la savane sans doute du fait qu'ils ne s'y sentent pas à l'abri.
En théorie, puisque nous sommes des hominidés, nous devrions découvrir des fossiles préhumains dans d'anciennes zones forestières. Le fait que nous ne trouvions pas de fossiles dans les paléoforêts peut s'expliquer par le fait que la nature du sol des forêts denses ne permet pas une bonne conservation des fossiles. Ce n'est probablement pas une coïncidence si les chercheurs n'ont pratiquement trouvé aucun fossile de chimpanzé ou de gorille; ils ont simplement vécu au mauvais endroit. Même s'il ne fait aucun doute que le chaînon manquant qui nous relie aux singes ainsi que les premiers homininés vivait dans les arbres - la forme de leurs pieds et de leurs mains le confirme -, les paléoanthropologues auront du mal et devront compter sur la chance s'ils veulent confirmer nos origines forestières. La "théorie de la Savane" semble donc être un cas particulier qu'il faudra un jour remplacer par une théorie plus générale expliquant pourquoi une espèce arboricole et en sécurité dans les arbres s'est mise soudainement debout sur les branches et trouva l'envie de fouler le sol, alors qu'elle savait d'instinct qu'elle risquait sa vie en y posant les pieds si d'aventure elle rencontrait des prédateurs (ce qui ne manquait pas d'arriver puisqu'ils passent leur temps à chasser en groupe ou à attendre leur proie à l'affût de jour comme de nuit). Les contraintes physiques Une autre cause de la bipédie serait une influence induite par les contraintes physiques. En effet, nous savons que des "gènes architectes" (par ex. la protéine Bmp4) tirent profit de lois de transformations géométriques, ce qui permet d'expliquer facilement la diversité des formes des espèces sans invoquer de mécanisme obscur. L'expression de ces gènes a des conséquences directes sur la morphologie, l'anatomie et donc également sur le squelette des animaux. Puisque le génome conserve la mémoire de l'adaptation de l'organisme dans son environnement, il est tout à fait possible que certaines espèces de vertébrés aient été influencées à la fois par la pression de l'environnement et par les effets induits par ces types de contraintes. Si nous pouvons isoler et identifier ces gènes et déterminer s'ils sont communs à tous les vertébrés et tous les préhumains, encore faut-il comprendre la manière dont ils s'expriment et leurs conséquences. Si la cartographie du génome humain a pris plus de 20 ans (1980-2001) et occupé des milliers de chercheurs ainsi que les plus puissants ordinateurs, déterminer le rôle individuel de chaque gène dans le détail y compris ses interactions et les effets d'une mutation (duplication par exemple) risque d'occuper les chercheurs bien plus longtemps. Dans tous les cas, au fil de l'évolution, par sélection naturelle ces influences ont pu induire des mutations génétiques qui eurent pour effet de modifier le rôle du cervelet, siège du contrôle moteur (postures et contrôle du corps) chez tous les vertébrés, en lui ajoutant de nouvelles capacités motrices et cognitives afin que son propriétaire appréhende mieux son environnement et survive dans de meilleures conditions. Le cervelet ne pouvant assurer seul toutes ces fonctions, le cortex cérébral pris la relève pour les fonctions supérieures. Malheureusement l'arbre phylogénique est trop incomplet et les fossiles trop épars pour proposer une explication définitive et une meilleure théorie. En revanche, si nous ignorons en partie pourquoi nos ancêtres se sont redressés, nous savons quels sont les avantages que procure la bipédie. Les avantages de la bipédie Nos ancêtres se sont redressés sur leurs pieds il y a plusieurs millions d'années pour ne plus jamais courber l'échine, même devant l'ennemi. Quel avantage procure la bipédie ?
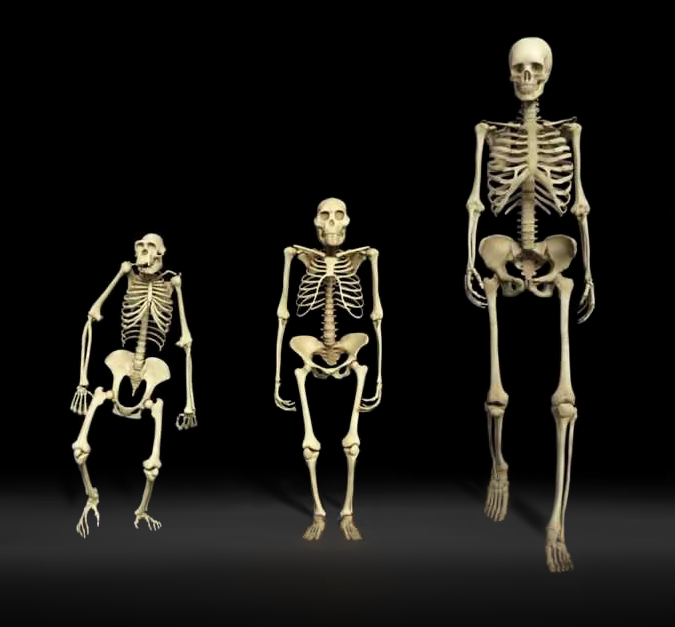
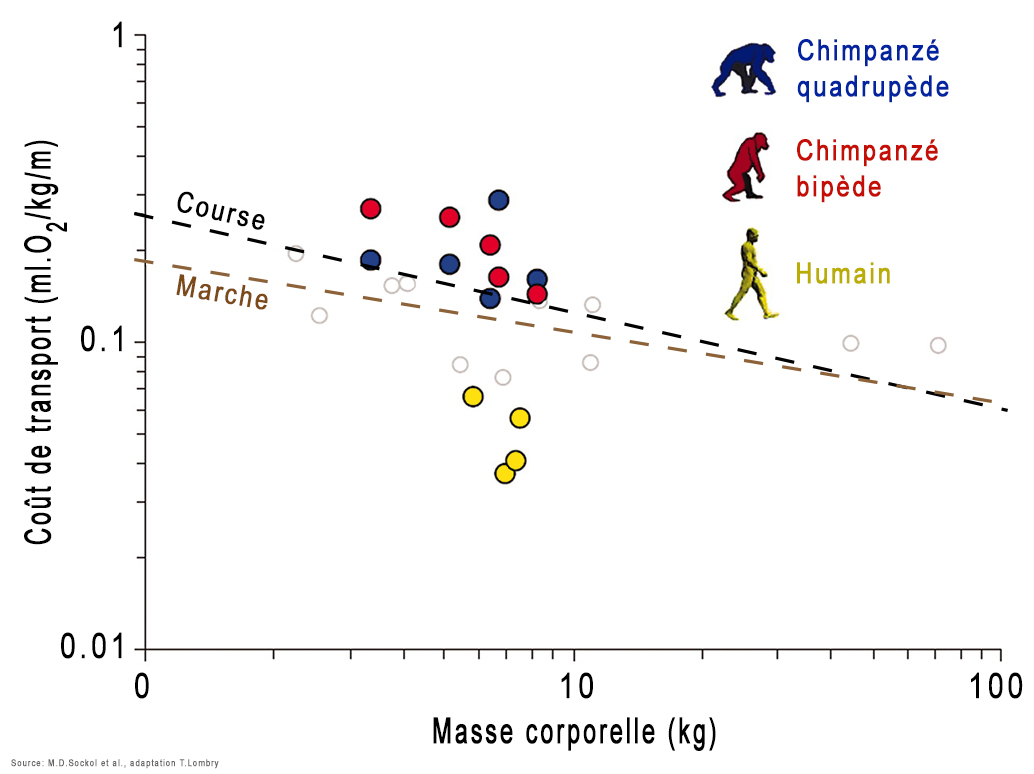
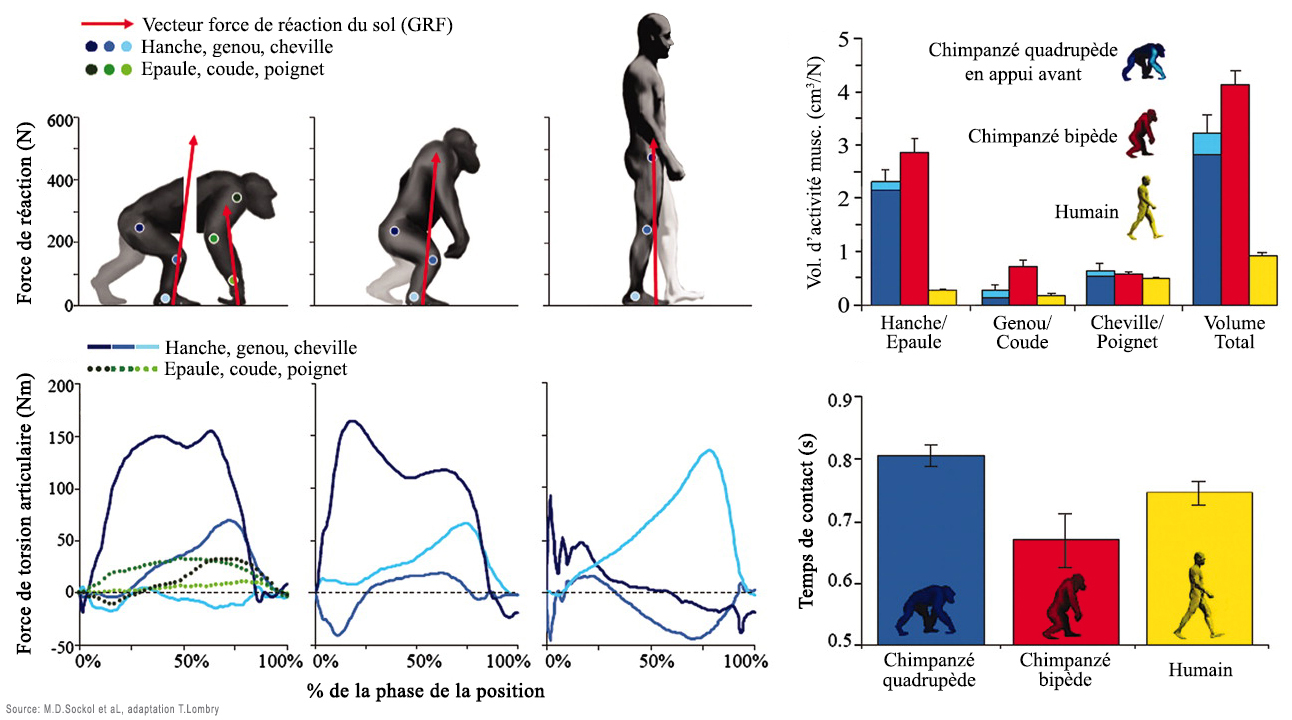
Comme on le voit à droite, la morphologie de Lucy (au centre de l'image) est beaucoup plus proche de celle de l'homme moderne que du chimpanzé. Rien que la forme des pieds de Lucy suggère qu'elle pouvait marcher debout. Un animal vivant à quatre pattes n'a aucun intérêt à se redresser car sa morphologie l'empêche d'en tirer profit; la position de son crâne le force à regarder le sol et la forme de son bassin entrave sa marche. A l'image des grands singes, il peut éventuellement se redresser s'il transporte des objets ou un bébé mais uniquement sur de courtes distances car sa démarche est déhanchée et donc fatiguante et mal assurée. De plus quand il essaye de marcher, ses fémurs sont très inclinés parfois presque à l'horizontal, créant à la longue des douleurs musculaires ainsi qu'au bas du dos. Aussi, pour marcher debout il faut que le squelette et les muscles supportent ce redressement de manière optimale, équilibrée et sans perte d'énergie afin que tout l'organisme en tire profit. Des mesures faites en laboratoire ont montré que la consommation d'énergie d'un chimpanzé marchant sur ses deux pattes arrière ou sur ses quatre pattes est presque identique. On peut donc imaginer que cette transition fut assez naturelle chez nos ancêtres arboricoles et n'exigea pas de gros efforts. En revanche, selon une étude réalisée par l'anthropologue Michael D. Sockol et ses collègues de l'Université de Californie à Davis, en marchant debout, l'homme a pu économiser jusqu'à 75% de l'énergie que consomme un chimpanzé marchant à quatre pattes ou debout et diviser par trois sinon davantage les forces de tensions articulaires. En étant capable de marcher debout, l'homme a pu utiliser l'excédant d'énergie pour développer son cerveau qui, rappelons-le consomme 25% de toute l'énergie que produit son corps, soit à lui seul environ 500 kcal./jour pour un homme moderne au repos. En parallèle, la station debout offrit à l'homme la possibilité de courir tout en transpirant et en économisant son énergie. Cette transformation de son anatomie et de sa physiologie ont offert au (super)prédateur qu'il est un avantage vital sur tous les autres animaux. En effet, l'analyse de l'horloge moléculaire des mutations de l'ADN a permis d'établir que les préhumains ont perdu leur pilosité il y a environ 3 millions d'années. La perte des poils permit aux hominidés d'améliorer leurs activités au quotidien, en se permettant notamment de chasser leurs proies en courant et même de travailler sous le Soleil des Tropiques pendant que les autres espèces récupéraient en faisant la sieste.
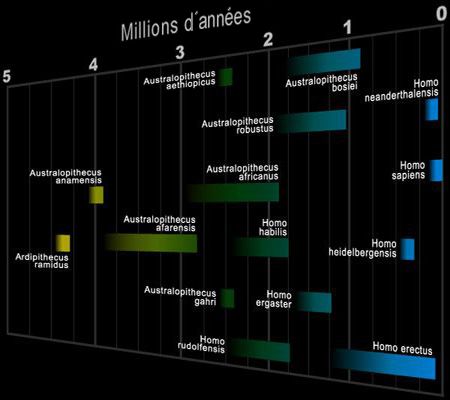
L'homme va transpirer par la peau (sa surface représente 1.7 m2 pour un homme de 1.70 m et 70 kg) en dissipant son excédant de chaleur au contact de l'air alors que les autres animaux ne peuvent dissiper leur chaleur corporelle que par la langue, le museau ou les oreilles. Aussi puissant ou rapide soit-il l'animal pris en chasse va s'épuiser et finir par s'immobiliser sous le coup de la chaleur. Ainsi encore aujourd'hui, les Bushmen peuvent chasser des proies agiles en courant quatre heures si nécessaire. C'est un avantage précieux quand la survie se conjuge au quotidien. En revanche, en perdant ses poils, l'homme a perdu son meilleur isolant face au froid; un animal couvert de fourrure ou de plume peut endurer l'hiver des pays nordiques sans précaution particulière (par ex. le chien husky, le cerf, le renne, le lynx, le léopard des neiges, l'ours blanc, le rouge-gorge, l'harfang des neiges, etc). En migrant vers le nord de la planète et les montagnes, l'homme dut s'adapter pour résister au froid et n'a pas eu d'autre choix que d'inventer la "peau de bête" chère aux Homo erectus et aux Néandertaliens. On y reviendra. Il y a quelques millions d'années de nombreuses espèces d'homininés foulaient la terre d'Afrique, ce qui soulève la question des raisons de cette multiplication du nombre d'espèces. La multiplication des espèces De Toumaï aux premiers homininés, soit sur une période de plus de 3 millions d'années, les paléoanthropologues ont découvert très peu d'espèces différentes et leur morphologie a peu évolué. En revanche, entre les Australopithèques (Lucy notamment) et Homo erectus et sur une période aussi étendue, le nombre d'espèces d'homininés a littéralement explosé et leur morphologie a fortement évolué, notamment la forme de leur crâne et le volume de leur cerveau. Comment expliquer ces changements ?
Pendant des décennies, les paléoanthropologues se sont demandés pour quelle raison nos ancêtres s'étaient mis debout et avaient conquis le monde. On a d'abord cru que nos ancêtres étaient descendus des arbres et s'étaient tout simplement adaptés à leur milieu, les différentes espèces étant apparues par hasard au gré des mutations graduelles ou ponctuelles. Si cette théorie explique beaucoup d'évènements, elle n'explique pas la prolifération soudaine des espèces il y a environ 3 millions d'années. En effet, cette diversification peu après l'époque de Lucy est en contradiction avec l'effet du hasard et nous devons chercher l'explication ailleurs, sans doute dans l'effet combiné de plusieurs phénomènes endogènes et exogènes. La paléontologie ne nous fournissant aucun indice, depuis quelques années les chercheurs se sont tournés vers d'autres sources d'informations. Nous avons vu comment la géologie, la paléobiologie et l'étude du paléoclimat notamment, nous ont apporté un nouvel éclairage et de nouvelles données sur cette période lointaine du passé de l'humanité. 1. Impact des changements climatiques sur les espèces Jusqu'aux années 1980, les chercheurs accordaient peu d'importance aux impacts du milieu et du climat sur l'évolution des préhumains; elles semblaient anodines mais il faut dire qu'on savait peu de chose du passé climatique et géologique de l'Afrique. On savait toutefois que la séparation de la lignée des humains de celle des grands singes s'était produite bien avant le Pléistocène, période qui débuta il y a 2.58 millions d'années durant laquelle la glace recouvrit de nombreuses régions d'altitude et durant laquelle la pluviosité était plus importante, tant par rapport à aujourd’hui que par comparaison avec la période précédent le Pliocène, il y a 5.33 millions d'années.
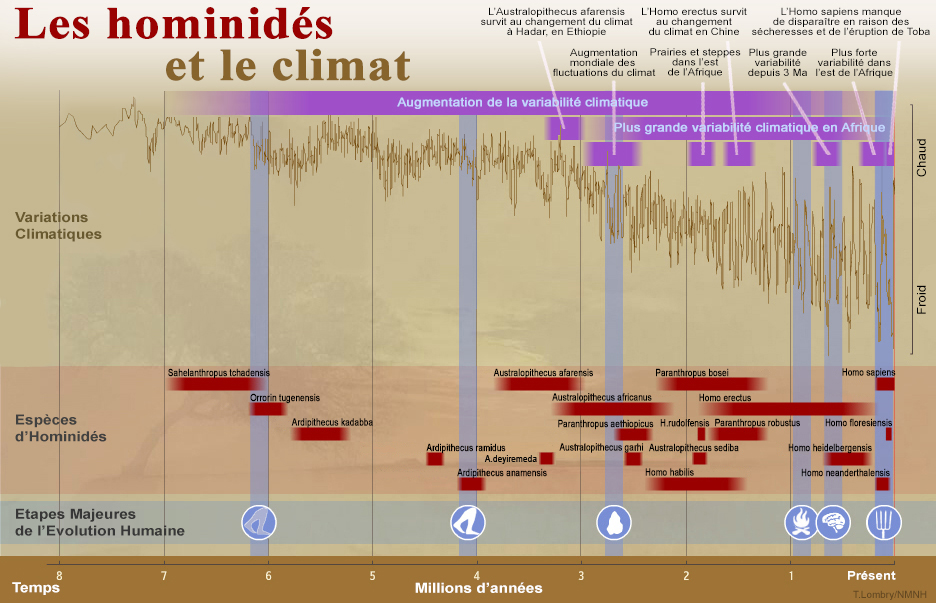
Entre la période où vécut Toumaï (7 Ma) et Lucy (3.2 Ma), soit pendant plusieurs millions d'années, le climat de l'Afrique est resté étonnamment stable. De ce fait, sans pression environnementale le nombre d'espèces d'hominidés et leurs caractéristiques n'ont pas subi de profondes modifications et par conséquent nos ancêtres n'ont pas inventé grand chose durant cette période. En analysant les couches géologiques du Rift ainsi que les couches sédimentaires du fond des océans, on a découvert qu'à l'époque de Lucy, les conditions climatiques ont sérieusement changé, le climat devenant beaucoup plus sec. En effet, à cette époque la Terre connut un “coup de chaleur”, les régions tropicales se sont asséchées et la grande forêt du Miocène devint herbeuse, se transformant graduellement en savane puis en prairie. Puis, il y a 140000 ans, la région du Rift connut de nombreuses périodes alternant forte humidité, y compris de grands lacs, et sécheresses qui ont duré plusieurs milliers d'années. Ces changements climatiques globaux et radicaux eurent des répercussions tant dans le règne animal que végétal. Un être vivant aussi complexe qu'un mammifère ou qu'un préhumain met des milliers d'années pour s'adapter à un changement climatique. Si son organisme est peu réactif, son espèce ne résistera pas longtemps et disparaîtra. En revanche, une espèce plus réactive, même moins spécialisée, sera avantagée et pourra s'adapter à ces changements. Nos ancêtres subirent cette évolution. Les ancêtres des homininés ne se sont donc pas simplement adaptés au milieu et au climat comme on le pensait jusqu'à présent. C'est le contraire qui s'est produit : le changement climatique fut le moteur de l'évolution des préhumains; c'est parce qu'il eut des changements climatiques répétitifs que nos ancêtres ont été contraints de développer leur corps et leur cerveau; ils devaient rapidement s'adapter ou périr. Le changement climatique explique pourquoi les préhumains se sont transformés : pour survivre, ils perdirent leurs canines et devinrent omnivores. Leurs canines se rétractant, ils perdirent leur prognathisme et leur visage s'ouvrit. L'ouverture de leur angle facial allait leur permettre d'articuler. Après être descendus des arbres dans lesquels ils s'étaient réfugiés durant des millions d'années, nos ancêtres se mirent debout. Toumaï comme Lucy étaient capables de marcher sur de courtes distances même si leurs pas étaient encore mal assurés. Puis nos ancêtres ont dû trouver de la nourriture par leurs propres moyens. Fragiles, glabres et sans défense, ils ont dû être plus intelligents, plus malins et plus habiles que les autres prédateurs. A défaut de courir vite, nos ancêtres étaient endurants, à défaut d'être puissants et de disposer de crocs ou de griffes ils étaient armés et chassaient en groupe. Aucun animal ne pouvait plus leur résister. L'homme devint un superprédateur. On a longtemps cru que les changements climatiques survenus au cours des 2 derniers millions d'années eurent un rôle clé dans l'évolution du genre Homo. Mais étant donné le nombre limité de données paléoclimatiques représentatives des régions d'intérêt anthropologique, il est resté difficile de quantifier ce lien. Dans un article publié dans la revue "Nature" en 2022, une équipe internationale de géophysiciens et de climatologues étudia les effets des variables astronomiques (excentricité et précession) et du climat sur les habitats des humains archaïques et l'évolution des espèces. Un résumé des résultats de leurs simulations est présenté ci-dessous.
Les chercheurs ont utilisé un modèle de la circulation générale atmosphérique au Pléistocène qu'ils ont combiné avec des données fossiles et archéologiques pour étudier l'adéquation spatio-temporelle de l'habitat pour cinq espèces d'homininés au cours des 2 derniers millions d'années. Ces espèces comprennent les Homo archaïques (H.ergaster et H.abilis), H.erectus, H.heidelbergensis, H.neanderthalensis et H.sapiens. Les chercheurs ont montré que les changements de la température, des précipitations et de la production primaire nette terrestre eurent un impact majeur sur la distribution géographique de ces espèces. Au cours du Pléistocène inférieur (2.58-0.78 Ma), les homininés se sont installés principalement dans des environnements à faible variabilité climatique, phénomène corrélé avec les paramètres orbitaux de la Terre. Ce comportement a considérablement changé après la transition survenue au milieu du Pléistocène, lorsque les humains archaïques ont quitté l'Afrique pour conquérir le monde et se sont adaptés à un large éventail de climats. Comme illustré ci-dessus, l'analyse du chevauchement simulé de l'habitat des homininés il y a environ 400-300 ka (l'époque de H.Heidelbergensis, H.Neanderthalensis et des premiers H.sapiens) suggère en outre que les perturbations climatiques antiphasées observées en Afrique australe et en Eurasie ont contribué à la transformation évolutive des populations d'Homo heidelbergensis respectivement en Homo sapiens et en Néandertaliens. 2. Impact des mutations génétiques Les découvertes de Toumaï, Orrorin, A.ramidus et Abel permettent aux paléoanthropologues de préciser l'évolution de la lignée des hominidés jusqu'à l'Homo sapiens. Tous les indices tendent à confirmer que l"adaptation sélective qui conduisit à ce dernier se déroula en Afrique où, à n'en pas douter, des milliers de gisements fossiles restent à découvrir près du Rift ou plus au nord-ouest, vers le Tchad. Mais son extension au reste du monde fait l'objet de beaucoup plus de spéculations. D'une part les paléoanthropologues sont frustrés du fait que les fossiles d'Australopithèques sont beaucoup mieux représentés que ceux des grands singes avec lesquels pourtant, ils sont contemporains. Nous avons vu que la présence des grands singes dans les forêts peut expliquer la disparition de leurs fossiles.
Si les chercheurs pouvaient trouver quelques fossiles de grands singes, ils comprendraient non seulement avec laquelle des deux espèces de Pan - aethiopicus, boisei, robustus d'un côté, paniscus, verus et troglodytes de l'autre - l'Homo sapiens moderne doit être comparé, mais cela apporterait également un nouvel éclairage sur les influences environnementales qui ont permis d'élaborer l'adaptation des deux lignées. Si le facteur climatique semble jouer un rôle clé dans l'évolution des espèces d'homininés, il faut aussi considérer que l’évolution des Australopithèques ne fut pas uniquement instiguée par des influences extérieures mais également par des avantages sélectifs provoqués par des mutations survenues dans le génome qui conféra à ceux qui en disposaient des avantages décisifs sur les espèces en compétition ou inadaptées. On a en effet découvert que l'Homo sapiens différait des grands singes par le fait qu’il disposait de 23 paires de chromosomes au lieu de 24. Les gènes des chromosomes manquants ont été identifiés comme formant un seul bloc localisé sur l'une des extrémités du chromosome 2. Il s'ensuit qu’à un stade du processus évolutionnaire, il y eut une translocation chromosomique qui incorpora l'entièreté d’un chromosome du singe dans le chromosome 2 humain. On ignore à quelle époque et pour quelle raison cette translocation s'est produite et quelles furent ses conséquences génétiques. Toutefois comme nous l'avons expliqué, d'après les analyses des mutations de l'ADN, l'époque de la divergence du genre Homo des chimpanzés remonte à environ 9 millions d'années. Cette translocation génétique pourrait expliquer le manque de corrélation dans l'histoire humaine entre les phénomènes d'adaptation et le milieu, bien que l'environnement dans lequel vécut l'Australopithèque soit très insuffisant pour porter ce jugement. Pour l'heure, nous en saurons peut-être un peu plus à ce sujet en étudiant le génome humain, en particulier en découvrant quel type de gène contient le chromosome 2. Nous savons que des erreurs de copie sont à l'origine de certaines maladies déjà identifiées et qu'un complexe de gènes HOX a été dupliqué au moins deux fois au cours de l'évolution pour des raisons encore inconnues. Tout ce qu'on peut dire avec certitude c’est que chaque espèce animale apporta sa solution au changement climatique : les animaux arboricoles devinrent fouisseurs, les homininés changèrent de dentition en changeant de régime alimentaire, leur cerveau passant de 400 à 800 cm3 environ en quelques centaines de milliers d'années. Alors que les espèces préhumaines subissaient une dernière transition ou hybridation génétique, l'espèce humaine apparut. Aussitôt que l'ancêtre de l'homme s'est mis à manger de la viande, il bougea, s'adaptant à l'environnement. L'évolution, sans nécessairement déterminer la nature et le devenir de l'homme a inscrit ce changement dans l'environnement qui a retenu les caractères dominants. Prochain chapitre
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
















{kind=link}