|
|
|
Origine et évolution de l'Homo sapiens
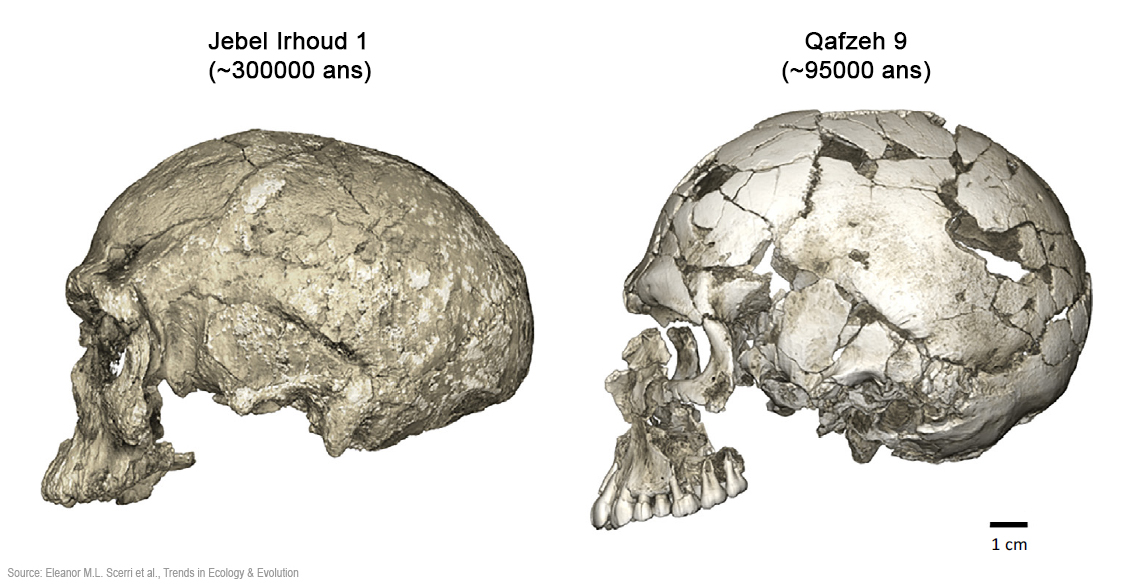
Vers une révision de l'évolution des Homo sapiens (I) Si notre origine africaine est établie, les différentes générations d'Homo sapiens descendent-elles d'une seule population ou d'un sous-groupe de cette population ? Et quelles sont les conséquences de l'une et de l'autre évolution ? En 2018, une équipe pluridisciplinaire composée de 23 chercheurs (généticiens, paléontologues, anthropologues, archéologues, zoologues et géographes) tenta de répondre à cette question dans un article publié en libre accès dans la revue "Trends in Ecology and Evolution" par l'éditeur de la revue "Cell" intitulé "Did Our Species Evolve in Subdivided Populations across Africa, and Why Does It Matter?" (Est-ce que notre espèce évolua en populations subdivisées à travers l'Afrique, et en quoi est-ce important ?). C'est la traduction intégrale - adaptée et non littérale - de cet article (et sans commentaires personnels) que nous vous proposons ci-dessous. En résumé Les chercheurs expliquent pourquoi ils contestent l'idée que l'espèce Homo sapiens aurait évolué au sein d'une même population et/ou région d'Afrique. En effet, les résultats des recherches montrent que les données pertinentes ne sont plus cohérentes avec cette vision. Fondant leur argumentation sur près d'une centaine d'études récentes, les chercheurs affirment que l'Homo sapiens évolua au sein d'un ensemble de groupes interconnectés vivant à travers l'Afrique, dont les relations évoluèrent au cours du temps, ce que les démographes appellent des connexions changeantes (cf. Simon Szreter, 1993, JSTOR, pp.659-701). Les modèles génétiques actuels doivent donc incorporer une vision plus complexe de la migration des populations à travers l'Afrique. Sur le plan paléontologique, la chronologie et la diversité physique des fossiles humains du Pléistocène (s'étendant entre 2.58 millions d'années et 11700 ans d'ici) suggèrent que des populations morphologiquement variées appartenant au clade Homo sapiens vivaient dans toute l'Afrique. De même, sur le plan archéologique les données provenant d'Afrique démontrent l'origine polycentrique et la persistance de cultures lithiques régionales distinctes au Pléistocène présentant une variété de paramètres paléoécologiques. Les études génétiques indiquent également que la structure des populations africaines tire ses racines loin dans le temps, parallèlement à un paléoenvironnement constitué de zones habitables mobiles et morcellées. Les chercheurs soutiennent que ces données supportent la théorie émergente d'une préhistoire africaine très structurée qui devrait être prise en compte dans le schéma évolutionnaire humain. Cette thèse devrait inciter les chercheurs à proposer de nouvelles interprétations et questions, les auteurs proposant également de nouvelles voies de recherches interdisciplinaires. Voyons cette thèse en détails. A consulter : Glossaire 1. Une vision différente de nos origines africaines La lignée des Homo sapiens est probablement apparue en Afrique il y a au moins 500000 ans[1] tandis que les premières manifestations morphologiques observées dans ce clade remontent à ~315000 ans[2]. Les premiers fossiles d'Homo sapiens ne présentent pas une simple progression linéaire vers la morphologie humaine contemporaine. Au lieu de cela, les premiers restes présumés d'Homo sapiens présentent une remarquable diversité morphologique et répartition géographique. Combinés aux récents gisements archéologiques et auyx nouvelles lignées génétiques, ces données sont compatibles avec le point de vue selon lequel notre espèce est originaire et s'est diversifiée au sein de populations fortement subdivisées (c'est-à-dire structurées), vivant probablement dans toute l'Afrique et étaient génétiquement reliées par un flux de gènes sporadique[1], [3], [4], [5], [6], [7], [8]. Ce concept de "multirégionalisme africain"[1] peut également inclure l'hybridation entre l'Homo sapiens et des homininés plus divergents vivant dans différentes régions[1], [9], [10], [11], [12]. Fondamentalement, de telles subdivisions de population seraient apparues et auraient survécu grâce à des changements des limites écologiques[7], [13], [14]. Cette hypothèse remet en question l'idée que notre espèce était endémique à une seule région (le Rift) ou à un seul habitat (la savane), impliquant l'existence d'une complexité souvent méconnue de nos origines africaines. Les auteurs ont ensuite fait la synthèse des données fossiles, archéologiques, génétiques et paléoenvironnementales pour essayer d'affiner notre compréhension de nos originaires africaines ainsi que les caractéristiques et la manière dont s'est maintenue la structure de la population du Pléistocène. Il fallut ensuite tenter de distinguer les données factuelles des erreurs et autre inférences sachant que l'utilisation de modèles informatiques simulant l'évolution de la structure d'une population modifie sensiblement les interprétations concernant l'évolution humaine récente. 2. La diversité morphologique et la propagation du clade Homo sapiens L'éventail des caractéristiques morphologiques caractérisant l'Homo sapiens ont fortement influencé les interprétations récentes sur les origines humaines, incluant ou excluant différents fossiles des interprétations. Par exemple, différents critères morphologiques et autant de méthodes analytiques ont été utilisées pour soutenir à la fois l'existence chez notre espèce d'un processus progressif de type mosaïque ou inversement, une spéciation ponctuée (par ex. [1]). Le crâne humain actuel est caractérisé par une combinaison de détails qui nous distinguent de nos parents fossilisés et de nos ancêtres, tels qu'un petit visage gracile, un menton et un cerveau globuleux. Cependant, ces caractéristiques humaines modernes typiques ont émergé à la manière mosaïque au sein du clade Homo sapiens. Comme on le voit ci-dessous à gauche, les plus anciens membres actuellement connus du clade Homo sapiens sont les fossiles découverts à Jebel Irhoud en Afrique du Nord datant d'environ 315000 ans qui présentent une morphologie faciale très similaire à celle de l'Homo sapiens ainsi que des volumes endocrâniens qui s'ajustent dans la gamme des variations contemporaines[2]. Cependant, la forme endocrânienne est allongée plutôt que globulaire, ce qui suggère que les caractéristiques distinctives endocrâniennes et probablement de la fonction cognitive ont évolué au sein de la population des Homo sapiens[2], [5].
D'autres anciens fossiles d'Homo sapiens excavés à Florisbad en Afrique du Sud (259000 ans), à Omo Kibish (~195000 ans) et Herto (~160000 ans), tous deux en Éthiopie, sont morphologiquement diversifiés[1], [16]. Cette diversité a conduit certains chercheurs à suggérer que les fossiles tels ceux de Jebel Irhoud et Florisbad représentent en fait une espèce plus primitive appelée "Homo helmei" sur base du nom du crâne partiel découvert à Florisbad en 1932[17], [18]. Dans la même catégorie, le crâne fossile de l'homme de Herto[19]qui combine un crâne relativement globulaire avec un os occipital robuste et une large face a été décrit comme la sous-espèce Homo sapiens idaltu du fait qu'il tombe en dehors de la variabilité des Homo sapiens récents[15]. Cependant, les chercheurs considèrent que l'Homo sapiens est une lignée en évolution ayant des racines africaines profondes et par conséquent préfèrent considérer ces fossiles comme faisant partie de la diversité initiée par les premiers membres du clade Homo sapiens. La série complète de caractéristiques crâniennes caractérisant les humains contemporains n'apparaissent qu'assez récemment, entre environ 100000-40000 ans[20]. Les caractéristiques et la chronologie des premiers fossiles d'Homo sapiens ainsi que leur répartition géographique à travers l'Afrique suggèrent que l'évolution a parfois progressé indépendamment dans différentes régions, dans des populations souvent à moitié isolées depuis des millénaires par la distance et/ou des barrières écologiques, comme des régions hyperarides (des déserts) ou des forêts tropicales. D'autres aspects de l'étendue géographique et de la diversité potentielle de l'habitat des premières populations d'Homo sapiens peuvent être obtenus en examinant des populations africaines de cueilleurs plus récentes qui étaient également fortement structurées. Par exemple, les restes humains de la fin du Mésolithique (anciennement la fin de l'Âge de la pierre moyen) à la fin du Pléistocène, il y a 20000 à 11700 ans mettent en évidence à la fois la conservation des caractères "archaïques" et le maintien d'une diversité morphologique importante[11], [21].
Pendant l'Holocène (depuis 11700 ans), la quantité de squelettes devient beaucoup plus importante, mais il reste aussi une importante variabilité morphologique spatiale. La variabilité entre les populations des différentes régions et environnements africains peuvent être le résultat de l'isolement dû à la distance et des adaptations environnementales locales[22], [23], [24], [25], [26]. Par exemple, des environnements difficiles (comme les déserts ou la forêt pluviale) et l'isolement ont probablement joué un rôle important dans la structuration de la population de chasseurs-cueilleurs africains de l'Holocène et des chasseurs-cueilleurs isolés dans la région des Tropiques[25], [26], [27]. En fin de compte, les processus expliquant l'émergence de tout "ensemble" de fonctionnalités à partir des résultats d'analyses de l'anatomie des premiers Homo sapiens restent incomplets et mal compris. Cependant, les données ne semblent pas correspondre à l'opinion admise de longue date selon laquelle la lignée humaine proviendrait principalement d'une seule région africaine accueillant une population homogène, dite panmictique. Au lieu de cela, l'Homo sapiens descendrait probablement d'une population structurée changeante (c'est-à-dire, un ensemble de groupes interconnectés dont les relations génétiques ont évolué), chaque groupe présentant des caractéristiques anatomiques de "modernité" différentes. La découverte de l'Homo naledi (~335000-236000 ans[28]) qui présente un aspect primitif et du crâne de l'Homo heidelbergensis de Broken Hill 1 en Zambie (~300000-125000 ans[29]) montre également que d'autres espèces d'homininés coexistaient en Afrique avec l'Homo sapiens, ce qui offre une possibilité de métissage archaïque sur le sol africain. Les recherches futures devraient essayer de déterminer quelles caractéristiques ont évolué avant l'apparition de notre espèce et lesquelles se sont principalement développées au cours de l'histoire évolutive de notre espèce. Un autre domaine clé concerne l'identification et la compréhension des différents processus à l'origine des changements observés. Par exemple, le rétrécissement du bassin peut refléter différents processus, y compris une dérive génétique neutre, l'adaptation aux changements écologiques et la variabilité de l'histoire du vivant. 3. Un patchwork culturel panafricain Partout en Afrique, l'abandon virtuel des grands outils de taille tels que les bifaces acheuléens au profit de la méthode de débitage Levallois (au Paléolithique moyen avec la fabrication de pointes minces et autres lames) beaucoup plus soignée marqua un profond changement technologique et stratégique de la culture lithique des homininés. Ces changements technologiques qui définissent la transition du Mésolithique se seraient produits à travers toute l'Afrique à des époques généralement similaires. Par exemple il y a ~300000 ans à la fois à Jebel Irhoud où on trouve les premiers fossiles d'Homo sapiens[16], à Olorgesailie en Afrique de l'Est[30]et il y a ~280000 ans dans le sud de l'Afrique à Florisbad[31]. Pour l'heure, les premières datations des fossiles découverts en Afrique de l'Ouest sont plus récentes, remontant à 180000 ans mais la région reste très mal caractérisée[32]. Le Mésolithique est associé à des fossiles d'Homo sapiens fossiles mais l'Homo naledi et l'Homo heidelbergensis ont probablement survécu jusqu'à la fin du Pléistocène (il y a 11700 ans). Des styles de cultures lithiques clairement régionaux, impliquant généralement des pierres taillées complexes ont émergé au Mésolithique. Par exemple, le Mésolithique comprend des bifaces lourds, des bifaces lancéolates, des éclats, des fragments, des lames, etc, probablement depuis au moins la fin du Pléistocène moyen, il a 126000 ans[33]. À la fin du Pléistocène (ou Pléistocène supérieur, il y a 126000 à 11700 ans), l'expansion des prairies et des savanes en Afrique du Nord engendra une occupation humaine dense associée à des caractéristiques telles que des technologies locales spécifiques comme les pointes de flèches à pédoncule (ou épaulement) présentées parmi l'éventail d'artefacts présenté plus haut[34]. À peu près au même moment, on constate l'émergence d'industries lithiques comparables dans certaines parties de l'Afrique australe. Comme en Afrique du Nord, certaines de ces industries sont également associées à d'autres aspects d'une culture lithique complexe comme l'usage de l'ocre, d'outils en os, de perles de coquillages et de gravures abstraites[35].
Une telle régionalisation est typiquement liée à l'émergence de facultés ou connaissances "modernes". Cependant, elle peut également refléter l'interaction entre les variables démographiques (par exemple la densité de population[36], [37], [38]) et les traditions savantes des sous-populations ou des dèmes comme semble le démontrer les différentes industries lithiques. Ainsi, le nord et le sud de l'Afrique, en plus d'être géographiquement éloignés, ont également été séparés par des facteurs environnementaux en raison de l'expansion et la contraction des forêts en Afrique équatoriale, phénomène synchrone avec l'amélioration des conditions climatiques en Afrique du Nord. D'autres facteurs, tels que la variabilité de l'habitat et l'adaptation aux conditions environnementales locales sont également susceptibles de jouer un certain rôle dans la diversification de la culture lithique. Bien que les différences géographiques soient claires à l'échelle continentale, les schémas spatiaux localisés sont plus difficiles à discerner. Les similitudes entre régions peuvent résulter d'un contact occasionnel entre populations ou par une adaptation convergente à des conditions environnementales communes. En Afrique de l'Est pour exemple, bien qu'il y ait vraisemblablement une certaine variabilité, il semble y avoir une continuité de la culture lithique dans la plupart des sites datant du Mésolithique (par exemple[39]). Dans de nombreuses régions du Mésolithique "générique" les groupes qui ne portent pas de signes particuliers de régionalisme sont communs[40].
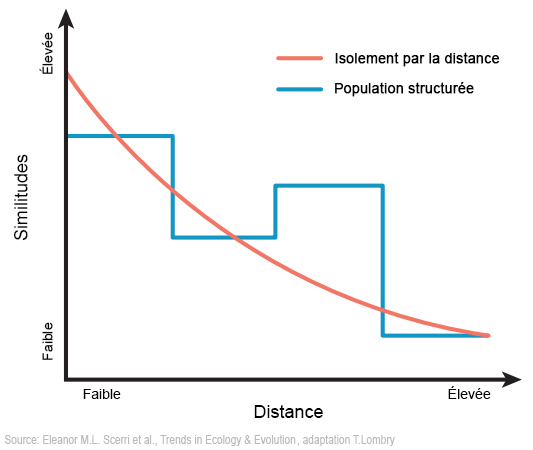
Placées dans un modèle cognitif, ces différences suggèrent que ces primo populations n'ont pas toutes présenté les signes d'un "esprit moderne". Cependant, de tels groupes sont plus nombreux du fait que la fréquence des changements des styles d'outils lithiques est plus élevée et pourrait représenter un indicateur spatial ou temporel reflétant probablement des facteurs démographiques. Dans certaines régions d'Afrique, l'éventail complet des caractéristiques du Mésolithique restent largement inchangées jusqu'à la limite Pléistocène/Holocène[41] et correspond aux schémas morphologiques, suggérant que la fin du Mésolithique put être aussi structurée et mosaïque que le début de la période. Ce point de vue est soutenu par la culture du Néolithique (qui s'étendit voici 60000 à 5000 ans). Malgré des similitudes superficielles visibles dans l'industrie microlithique du Néolithique, les données culturelles montrent une différenciation continue et une dérive durant l'Holocène qui viennent appuyer les preuves biologiques relatives à la dynamique de la variabilité des populations qui n'a pas abouti à une homogénéisation à grande échelle[42]. Les raisons et donc les implications de la structuration géographique et temporelle de la diversité culturelle au Mésolithique est encore mal caractérisée et reflète probablement plusieurs processus comprenant les adaptations à différents environnements[43]. À long terme et à grande échelle, la séparation des populations peut également avoir été la norme pour une grande partie de l'Afrique du Pléistocène (c'est-à-dire suite à l'isolement par la distance et l'isolement par l'habitat qui représentent des modèles nuls à rejeter, cf. Encadré 1 plus haut). Les rares modèles spatialement explicites explorant les innovations technologiques du Pléistocène ont également établi une corrélation entre la complexité culturelle et la variabilité des schémas régionaux de croissance, de mobilité et d'interdépendance de la population (par exemple [36], [44], [45]), supportés par des preuves du transfert à longue distance de la matière brute de l'industrie lithique (par ex.[46]). On peut suggérer que les nouvelles directions majeures de la recherche archéologique devraient : (i) démêler les contributions relatives des différentes régions / habitats africains à l'évolution humaine récente; (ii) comprendre les dérives des schémas de la structure des populations à travers l'apparence différentielle, l'expansion, la contraction et la disparition des formes distinctes d'artefacts régionaux (cf. Encadré 1); (iii) exploiter les liens croissants entre l'archéologie, l'écologie, la morphologie et la génétique pour explorer dans quelle mesure les schémas de la culture lithique sont couplés ou découplés de ceux qui lui sont associés (mais potentiellement indépendants). Prochain chapitre 4. La nécessité d'une vision plus complexe
|
||||||||||||||||||||||||