|
|
|
L'évolution des systèmes vivants
La génétique des populations (IV) Le domaine bien particulier de la génétique des populations a été fondé indépendamment par Sewall Wright, John Burdon S. Haldane et plusieurs généticiens. Ils fondèrent leurs arguments sur le fait que lorsque qu’une mutation directe survient et si elle est favorable pour l’individu, son extension ultérieure dans la population dépend de plusieurs variables : - La taille de la population - La longévité des générations - Le degré de viabilité de la mutation - Le taux avec lequel la même mutation réapparaît dans la descendance. Nous avons vu à propos des origines de la vie qu’un gène donné est favorable sous certaines conditions environnementales. Si celles-ci changent dans l’espace, il se peut que le gène ne soit favorable que pour une partie seulement de la population; si les conditions évoluent dans le temps, en général le gène devient inefficace. Etant donné que tous les individus contiennent habituellement un assortiment particulier de gènes, à l’exception des jumeaux homozygotes, le nombre total de gènes disponible pour la prochaine génération constitue un vaste réservoir potentiel de variables génétiques, ce qu’on appelle un “pool génétique”. Les virus en sont le plus bel exemple.
La reproduction garantit que les gènes seront réarrangés à chaque génération dans le processus de recombinaison. Dans une population stable, la fréquence avec laquelle un gène réapparaît est proportionnelle au nombre total de gènes dans le pool génétique; elle reste constante même si les gènes se recombinent différemment dans chaque individu. En revanche, si la fréquence génétique change d’une façon sensible, nous assistons à une évolution. C’est ainsi que les mutations offrent une possibilité au pool génétique d’être réalimenté en nouveaux gènes. Ce processus complète la sélection naturelle qui, en modifiant la fréquence génétique donne aux gènes avantagés plus de chances de se reproduire. Bien que cette théorie soit supportée par des lois mathématiques, elle ne sera pas reconnue jusqu’à la fin des années 1930, époque à laquelle Theodosius Dobzhansky vérifia les arguments proposés en laboratoire et par l’observation du milieu. Il démontra ainsi que l’adaptation génétique évoluait dans les grandes populations de mouche des fruits et résultait d’un changement environnemental contrôlé. Dobzhansky prouva que les observations génétiques étaient compatibles avec la sélection naturelle de Darwin, qui était à la source des plus petites modifications de la fréquence génétique et donc des changements évolutionnaires des caractères d’une population. Rappelons qu'en général un hybride naturel né de parents de deux espèces différentes mais appartenant à la même famille est stérile à quelques exceptions près (on a par exemple enregistré quelque 60 naissances chez les mules depuis 1527). Il existe également des croisements féconds entre le monde animal et végétal (cf. le cas de la limace de mer Elysia chlorotica). Cette nouvelle façon de comprendre la théorie de Darwin revitalisa pratiquement tous les champs de la biologie et de la paléontologie. La théorie synthétique ou néodarwinisme
Ce n'est qu'en 1882 que l'on parla explicitement de la génétique en biologie. Le biologiste allemand August Weismann émit l'hypothèse que seule les cellules sexuelles étaient responsables de l'hérédité, alors que le reste de l'organisme n'était qu'une structure qui devait disparaître avec la mort de l'individu : “La sélection naturelle, disait Weismann [15] n’opère en apparence que sur les qualités de l’organisme adulte, mais en réalité elle opère sur les dispositions cachées dans la cellule germinale. Tous les changements dus à des influences extérieures, sont de nature passagère et disparaissent avec l’individu [...]. L’essence de l’hérédité est la transmission d’une substance nucléaire d’une structure moléculaire spécifique”. A ces yeux, les cellules sexuelles (germen) étaient potentiellement immortelles. Il prouva sa théorie en coupant la queue de plusieurs générations de souris pour constater que cette mutilation n'avait aucun caractère héréditaire. Grâce aux idées de Weismann, nous distinguons aujourd'hui deux ensembles de caractères chez l'individu : le génotype, qui caractérise l'ensemble de son matériel génétique, et le phénotype qui caractérise l'apparence de l'individu. Par la suite, Grégor Mendel confirmera en 1860 que les caractères héréditaires se répartissaient de façon précise chez les individus, mettant un terme à la théorie statistique de Karl Pearson et ses amis. On retrouvera alors la loi de la sélection naturelle et la théorie génétique de Weismann. En faisant l'hypothèse qu'un gène devait être présent ou absent pour produire un effet héréditaire, Mendel supposait que l'apparition d'une nouvelle fonction devait se faire d'un seul coup et non pas graduellement comme Darwin l'imaginait. De Vries confirma expérimentalement la théorie de Mendel en 1886, en observant des mutations brusques sur des plantes mises en culture. Lamarckistes et Darwiniens contestèrent cette théorie mais ce fut en vain. L'expérience de la couleur des yeux démontrait qu'il n'y avait jamais d'enfant dont les yeux avaient des couleurs intermédiaires. Mais il y avait des contre-exemples, juste ce qui fallait pour réfuter la théorie : la variabilité de la taille, la couleur des cheveux ou de la peau, la prédisposition de certains à l'embonpoint, etc. A
voir : Les
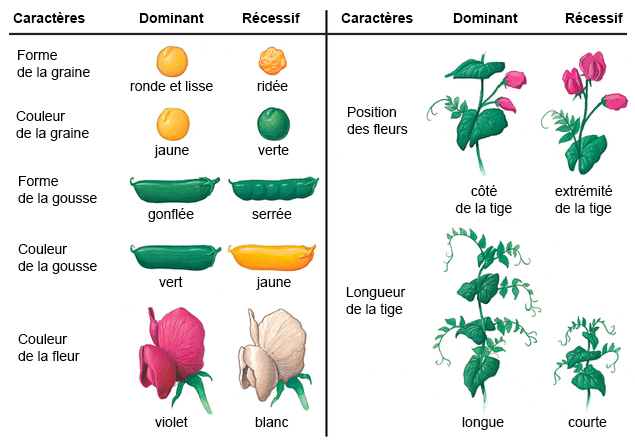
lois de Mendel
En 1918, le biologiste et biométricien Ronald A. Fisher[16] découvrit qu'il était possible de créer à la deuxième génération d'une variété des hybrides de variation continue : la couleur du blé ou des tulipes pouvait passer graduellement d'une couleur à l'autre. Plus tard, le zoologiste Ernst Mayr démontra à son tour que les nouvelles espèces apparaissaient en général dans un lieu géographiquement isolé et subissaient une véritable “révolution” génétique qui transformait rapidement le contenu de leur pool génétique. Le botaniste G.Ledyard Stebbins démontra enfin que les plantes se développaient selon un schéma comparable à celui des animaux. Il démontra en particulier que leur réponse adaptatives était liée aux pressions environnementales et fonction des opportunités. Les découvertes de Fisher, Mayr et Stebbins s’appliquaient ainsi à un grand nombre de systèmes vivants dont les preuves systématiques donnèrent naissance à une nouvelle théorie qui réconciliait enfin Mendel et Darwin : la théorie synthétique de l'évolution ou néodarwinisme qui servit de fondement à la théorie évolutionnaire à partir des années 1950. Darwin y retrouvait certaines de ces idées : sélection naturelle, adaptation des individus et des populations, les mutations dues au hasard. La science moderne y a ajouté la distinction entre les gènes et les caractères ainsi que les découvertes de la génétique moléculaire, ce dont Lamarck n'avait vraiment pas envisagé, lui qui considérait l'évolution comme l'hérédité des caractères acquis.
Aujourd'hui la théorie de l'hérédité des caractères acquis est oubliée. Nous savons que les gènes sont à la base de l'évolution embryonnaire. Or ceux-ci ne constituent pas un plan détaillé de l'individu à naître. Le développement d'un organisme est un processus qui rejoint plus l'idée d'une recette constituée de plats différents et formant une composition, qu'un plan détaillé de l'ensemble. Christian de Duve[17] résume cette idée dans le titre d’un chapitre qu'il consacre à la chimie prébiotique : "Construire sans plan". Prenons une fois encore l'exemple de la callosité de la voûte plantaire souvent citée. A force de marcher pieds nus, il est exact que la plante des pieds va s'endurcir à force de subir des actions mécaniques. En réalité, si ce phénomène existe, ce n'est pas l'hérédité des caractères qu'il faut invoquer. Il n'existe en effet pas de gène de la callosité de la voûte plantaire ou des doigts de la main, voire de l'apprentissage ! Les biologistes et les intellectuels qui raisonnent ainsi pensent implicitement qu'il existe une embryologie "préformiste" qui serait à tous les coups bénéfiques pour l'individu. Mais peut-on réellement soutenir que les caractères acquis sont toujours positifs ? Durant sa vie, un organisme subit de nombreux avatars, des accidents, des mutilations parfois. Pourquoi ces accidents ne seraient-ils pas eux aussi transmis aux enfants, sans parler des problèmes liés à la vieillesse... Non, la seule théorie rationnelle qui explique tout à la fois la callosité de la voûte plantaire et de façon générale l'adaptation des individus est la sélection naturelle.
La théorie synthétique de l'évolution cache malgré tout des zones d'ombres. On n'explique toujours pas l'existence des animaux aveugles en termes d'influence environnementale ou de survie du plus apte, alors qu'on a tendance à expliquer la morphologie des pingouins par l'usage. Et que dire du squelette "hybride" de l'ornithorynque ou de la survivance au fil de l'évolution du Coelacanthe ou de la prêle ? Pour en tenir compte, la théorie de l'évolution de Darwin doit être amendée. Prochain chapitre Les équilibres ponctués et les périodes de stase
|
|||||||||||||||||||||||||||||||||||||||||











{kind=link}